اندونوکلئازهای مهندسی ژنتیک به چهار گروه تقسیم میشود: هستههای شیمیایی، ZFNها، مگانوکلئاز و TALEنوکلئاز. این بخش به مگانوکلئازها اختصاص داده شده است که بیش از ۱۵ سال برای تحریک هدف گیری ژنی استفاده شده است. پیشرفتهای اخیر در مهندسی مجدد ویژگیهای مگانوکلئازها، دامنه کاربرد آنها را نیز افزایش داده است. نمرات نشریات و مشارکت در جلسات علمی مربوط به مگانوکلئاز، نشان دهندهی سیر تکاملی در پذیرش و کاربرد آن و یک جامعهی در حال رشد علاقهمند به خواص استثنایی این پروتئینها است.
خانواده مگانوکلئازها
مگانوکلئاز، اندونکلئاز هدفیاب نیز خوانده میشود که بر اساس توالی و ساختار عمده به پنج گروه تقسیم میشود: LAGLIDADG، GIY-YIG, HNH, His-Cys box و PD-(D/E)XK. گروه پروتئینهای LAGLIDADG بیش از همه مورد مطالعه قرار گرفتهاند و در همه قلمروهای زندگی یافت میشوند. عموما داخل اینترونها و intein ها یافت میشوند، گرچه در اعضای مستقل نیز یافت میشوند. تا به امروز، یک نقش هدفمند در میزبان برای این پروتئینها شناخته نشده است و تمایل دارند در گروه «عناصر ژنتیکی خودخواه» طبقهبندی شوند. با وجود چند استثنا پروتئینهای LAGLIDADG یک یا دو فعالیت اصلی را نمایش میدهند:
۱. آنها به عنوان یک RNA maturase در تسهیل اتصال اینترون خود عمل میکنند.
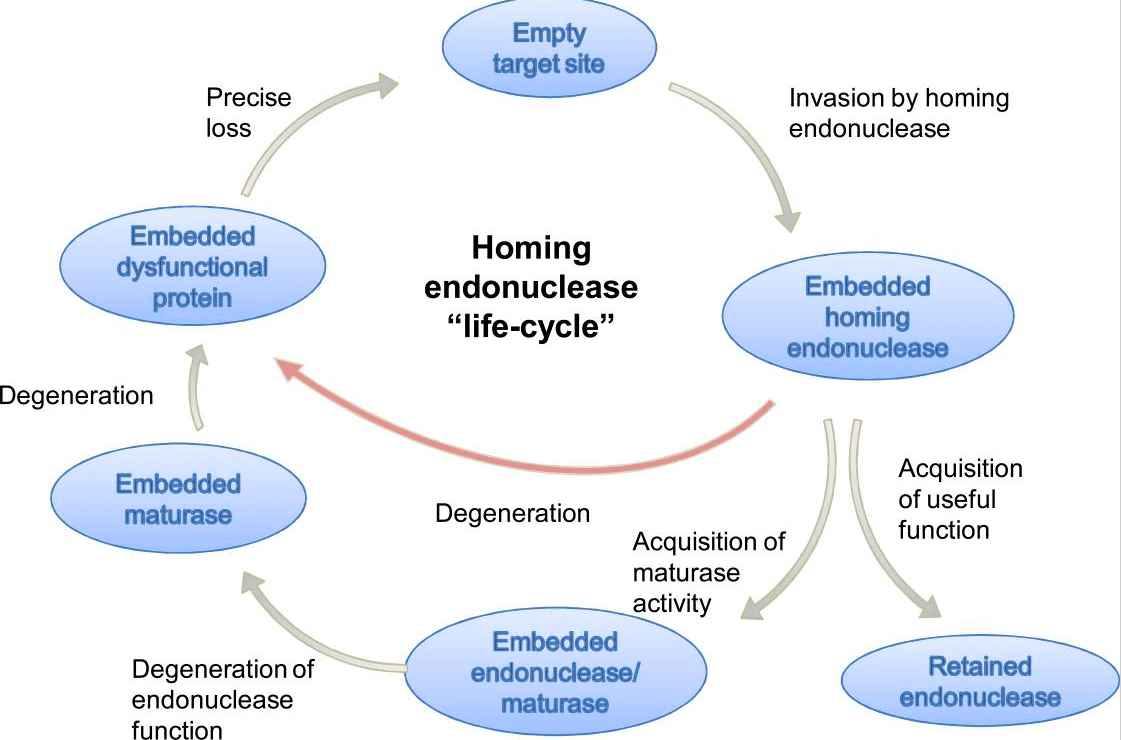
۲. هم چنین به عنوان اندونوکلئاز بسیار ویژه، توانایی تشخیص و شکافتن اتصال توالی اگزون-اگزون درحالیکه اینترون آن ثابت باشد را دارد. بدین ترتیب لقب «اندونوکلئاز هدفیاب» میگیرد. فرض بر این است که اندونوکلئاز هدفیاب یک چرخه زندگی مانند شکل زیر دارد:
(i) در وهله اول به عنوان اندونوکلئاز تهاجمی که توانایی تجهیز توالی کد شده را دارد شروع بکار میکند (ii) پس از تهاجم آنها به فعالیت یک RNA maturase همراه برای کمک به اطمینان از ترکیب مناسب اینترونشان نیاز دارند (iii) با گذشت زمان، فعالیت نوکلئاز مهاجم از بین میرود و تنها فعالیت RNA maturase باقی میماند و (iiii) در نهایت، با از دست دادن فعالیت maturase، تکثیر اینترون غیرممکن میشود و اینترون از بین میرود. بنابراین نتیجه گیری میشود که عملکرد یک پروتئین LAGLIDADG (اندونوکلئاز, maturase و یا هردو) یک تصویر لحظهای از وضعیت فعلی چرخه زندگیاش میدهد.
ساختار اندونوکلئاز LAGLIDADG
جسمی که هم از لحاظ عملکرد ژنتیکی و شیمیایی ثابت کرده است که اندونوکلئازهای هدفیاب LAGLIDADG میتوانند به عنوان ابزار مولکولی مورد استفاده قرار گیرند. مدت زیادی است شکل عمده توالی مشخص شدهی LAGLIDADG یک عنصر ضروری برای فعالیت آنزیمی است. برخی پروتئینها تنها فقط یک موتیف (شکل عمده) دارند، درحالیکه دیگران دو موتیف دارند: در هر دو مورد موتیف با ۷۵-۲۰۰ آمینواسید باقی مانده دارند تا هیچ شباهتی به دیگر اعضای خانواده نداشته باشند. در سال ۱۹۹۷، دو گروه سازمانبندی پروتئین LAGLIDADG را با ساختار دو پروتئین تک موتیفی (I-Crel) و یک پروتئین کدگذاری شدهی intein دابل موتیف (PI-Scel) روشن ساختند. در هردو مورد در حوزه اندونوکلئاز αββαββα، با موتیف LAGLIDADG متشکل از ناحیه ترمینال هلیکس اول جا میشود، که به یک مرکز کاتالیزوری دو طرفه کمک میکند، بلکه متشکل از تعامل زیرواحد/زیرواحد هسته است. دو حوزه چونα/β شکل گیری یک پروتئین را با رشتههایβ در هر منطقه شکل گیری ناحیه اتصال DNA زینی شکل، فراهم میاورند. زیرواحدهایی چون این دو که برای عملکرد نیازند باید به طور قطع ساخته شوند. همانطور که نشان داده شد، I-Crelهومودیمر، دارای معماری کلی مشابه حوزه دابل-موتیف اندونوکلئاز PI-Scel است. شکل α/β مشخصه پروتئین LAGLIDADG است و از آنجایی که برای مدل سازی همولوگ و پیش بینی ساختار/عملکرد پروتئینهایی که اطلاعات ساختاری ندارند، استفاده میشوند.
مقالات مرتبط: TALEN- افکتور نوکلئاز شبه فعال کننده رونویسی
برآورد شده که پروتئین LAGLIDADG به طور خاص یک توالی DNA در حدود ۱۴-۴۰ جفت پایه در درازا است. گرچه ساختار اولیه در غیاب DNA هدف حل شده، بسیاری از آزمایشگاهها موفق به استخراج اطلاعات ساختاری برای درک بهتر تعاملات پروتئین-DNA شدهاند. ابتدا، از ساختار PI-Scel روشن بود که حوزه اتصالی اضافه شده مشخصه پروتئینهای کدگذاری شده با intein در طرفین منطقه قادر به تماس مستقیم با DNA بود. زمانیکه ساختار کو-کریستال در نهایت حل شد، یک مفهوم ایجاد شد. این مطالعات، همراه سیر تکامل حوزه اندونوکلئاز (حوزه l) حوزههای اتصالی اطراف (حوزه I) را نشان میدهند. در وهله دوم، داشتن اطلاعات ساختاری، اجازه پیش بینی بهتر راجع به مطالعاتی که تلاش در بررسی عوامل تعیین کننده خاص دارد را میدهد. در حالیکه قبلا تصور بر این بود که اندونوکلئاز هدفیاب، قادر به تحمل تغییرات ژنتیک تک-پایه در طول توالی تشخیص آنها باشد و مطالعات انحطاط محل توالی میتواند تفسیر شود. ( I-Crel و PI-Scel) با این وجود، مدت کوتاهی بعد از آن ساختار کو-کریستال DNA در دسترس بودکه نه تنها نگاه اجمالی اولیه اتصال پروتئین DNAی LAGLIDADG، (I-Crel) را میدهد بلکه امکان مطرح شدن تئوری در رابطه با ماهیت کاتالیزوری را هم میدهد. علیرغم هر زیرواحد ساختار کو-کریستالی طبیعت ویژهی مخصوص را به تصویر کلی اثر متقابل پروتئین DNAی LAGLIDADG اضافه میکند. که بر چند اصل کلی پافشاری دارد: (i) تماسهای خاص از دفن رشته بتای گسترش یافته در شیار اصلی DNA، با ستیغ متصل سازی DNAی دارای اوج و حدفاصل که شبیهساز پیچ هلیکال DNA است (ii) در عین گستردگی، مکمل کامل پتانسیلی اتصال هیدروژنی بین پروتئین و DNA با تماسهای بسیار میانجی آب، هیچ وقت به طور کامل درک نمیشود (iii) برش برای تولید مشخصه ۴-nt 3’-OH برآمدگی در سراسر شیار کوچک اتفاق میافتد، درحالیکه پیوند scissile phosphat با تحریف در مرکز ناحیه «۴-base» به هسته کاتالیزوری پروتئین نزدیکتر میشود (iv) شکاف با یک مکانیزم طرح دو فلز اتفاق میافتد، گاهی اوقات یک نمونهی منحصر بفرد «به اشتراک گذاری فلز» را دربردارد. (V) و در نهایت افزودن پیوستگی و یا تماس مخصوص که میتواند از فرم «درست کردن» داربستها، در مناطق خارج حصار هسته α/β بوجود بیاید.
در بازبینی اول به نظر میرسد این کلیات میتواند مجموعهای از قوانین ثابت را تعریف کند که میتواند در کنترل تعاملات پروتئین LAGLIDADG كمك كند. با اين حال، مطالعات متعدد نشان دادهاند که پروتئینهای مرتبط میتوانند زیر مجموعههای مختلفی از باقی ماندهها برای تشخیص DNA مشابه استفاده کنند.