پتانسیل غشا چیست؟
هر واحد عملکردی عصبی به عنوان یک واحد الکتریکی عمل می کند و برای ایجاد جریان عصبی در مدار الکتریکی به اختلاف پتانسیل نیاز است؛ این اختلاف پتانسیل با وجود کانال های غشایی و ایجاد نفوذپذیری در غشا واحد های عصبی به وجود می آید که برای جا به جایی بار های الکتریکی (یون ها) در دوسوی غشا و ایجاد جریان عصبی برای انتقال پیام ها ضروری است.
در ایجاد اختلاف پتانسیل مذکور چند یون نقش اصلی را ایفا می کنند که عبارتند از : سدیم، پتاسیم، کلر و کلسیم. هر کدام از این یون ها بر اساس اختلاف غلظت و نفوذ پذیری خود اختلاف پتانسیل خاص خود را سبب می شوند و مجموع این اختلاف پتانسیل ها با هم جمع شده و اختلاف پتانسیل غشا را سبب می شوند.
اختلاف پتانسیل هر یون را با معادله نرنست (Nernst) اندازه می گیرند که به صورت زیر است :
(EMF)(mv)=61. log (In concentration/Ex concentration)
برای مثال اختلاف پتانسیلی که یون سدیم ایجاد می کند حدودا ۶۱- است؛ چون غلظت آن در بیرون سلول حدودا ۱۰ برابر غلظت آن در درون سلول است و لگاریتم ۰.۱ در مبنای ۱۰ براابر ۱- می شود . این عدد منفی به این معناست که غلظت در درون سلول کمتر است؛ چون در این معادله ما پتانسیل بیرون سلول را برابر صفر در نظر می گیریم.
پتانسیل آرامش چگونه محاسبه میشود؟
پتانسیل آرامش را با جمع هر کدام از پتانسیل های ذکر شده در بالا برای هر یون تاثیرگذار بدست می آوریم. با این تفاوت که باید نفوذ پذیری (permeability) هر یون را در معادله در غلظت آن ضرب کنیم. مثلا غشای سلول ها نفوذ پذیری خاصی برای یون های منفی مثل یون کلر ندارند ولی در عوض نسبت به یون پتاسیم بسیار نفوذ پذیرند.
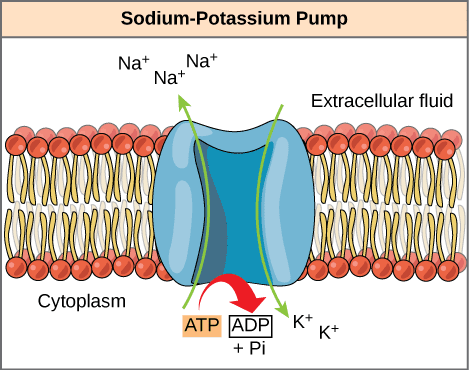
با جمع اعداد حاصله از معادله ها ۹۴- برای یون پتاسیم، ۸+ برای یون سدیم و ۴- برای فعالیت پمپ سدیم-پتاسیم در نظر گرفته می شود که با جمع این اعداد به عدد ۹۰- برای پتانسیل آرامش غشا می رسیم.
پمپ سدیم-پتاسیم به ازای هر ۳ یون سدیم که از سلول خارج می کند، فقط ۲ یون پتاسیم را وارد می کند و به این خاطر سبب منفی تر شدن سلول و حفظ پتانسیل آرامش می شود؛ چون هر چقدر درون سلول منفی تر، تحریک پذیری آن کمتر.
چه کانالهایی در انتقال یون ها نقش دارند؟
- پمپ سدیم-پتاسیم؛ همان طور که گفته شد سبب حفظ پتانسیل آرامش غشا می شود و در غشای همه سلول ها وجود دارد؛ چون این پتانسیل آرامش فقط مختص نورون ها نیست
- کانال های نشتی پتاسیم؛ این کانال ها دلیل نفوذ پذیری زیاد غشا ها به پتاسیم اند و مقدار کمی هم به سدیم اجازه عبور می دهند
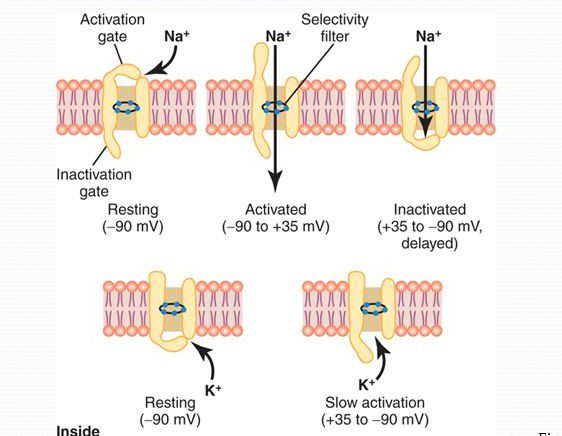
- کانال های دریچه دار (ولتاژی) سدیمی؛ دو دریچه در دو سطح خود دارند. دریچه ای که در سطح بیرون سلول قرار دارد با نام Activation gate شناخته می شود و در حالت آرامش بسته است و با بالا رفتن مقدار کم ولتاژ با تغییر شکل فضایی باز می شوند. دریچه رو به داخل این کانال ها با رسیدن ولتاژ به حد خاصی پس از شروع پتانسیل عمل باعث بسته شدن این کانال می شود. این کانال ها برخلاف نوع بعدی فعالیت سریعی دارند
- کانال های دریچه دار (ولتاژی) پتاسیمی؛ یک دریچه در سمت داخل دارند و به کندی عمل می کنند. این کندی عمل بالا رفتن ناگهانی پتانسیل غشا در ابتدای پتانسیل عمل و پایین آمدن تدریجی آن را توجیه می کند.

- کانال های دریچه دار کلسیمی؛ به مقدار کمتر و به ویژه در سلول های عضلانی وجود دارند. به میزان ۲۰ برابر کندتر از دریچه های سدیمی عمل می کنند. این کندی عمل به همراه کندی عملکرد کانال های پتاسیمی در سلول های قلبی، مسئول انقباض طولانی مدت این سلول ها هستند.
چرا کاهش غلظت کلسیم خارج سلولی خطرناک است؟
یون های کلسیم به پروتئین های کانال های سدیمی متصل می شوند و در شرلیط فیزیولوژیک با تحت تاثیر قرار دادن میدان الکتریکی آن ها ولتاژ لازم برای فعال شدندنشان را تغییرمی دهند. زمانی که مقدار یون های کلسیم کم شود نفوذ پذیری غشا به نورون ها زیاد شده و باعث ایجاد تحریک های خود به خودی می شود که باعث ایجاد تتانی می شود.
چرا نورونهای میلیندار و قطورتر ایمپالسها را سریعتر منتقل میکنند؟
برای پاسخ به این سوال باید کمی از “نظریه کابل های الکتریکی” اطلاع داشته باشیم. همان طور که وقتی از یک منبع صدا دورتر می شویم شدت صدای آن کاهش می یابد، جریان عصبی نیز هر چه از مرکز تولید خود دورتر می شود پتانسیلش کاهش می یابد. برای اندازه گیری این تغییر با دو ثابت سر و کار داریم:
- ثابت طول (lambda): چقدر مسافت لازم است تا ولتاژ یک جریان به صفر برسد.
- ثابت زمان (T): چقدر زمان لازم است تا ولتاژ یک جریان به صفر برسد.
- Cm: گنجایش غشایی کمیتی برای برای اندازه گیری ظرفیت یک غشا برای ذخیره بار الکتریکی است
با این توصیفات یک نورون ایده آل ثابت طول بی نهایت و ثابت زمان صفر دارد!
این دو ثابت، غیر فعال اند و زمانی فعال می شوند که کانال های یونی به کار بیفتند و جریان را تقویت کنند. هر بار که جریان به این شیوه تقویت شود حدودms 1 در آن تاخیر به وجود می آید و هر چقدر ثابت طول نورون کمتر باشد تعداد این تاخیر ها نیز بیشتر می شود؛ پس برای افزایش ثابت طول چه کاری می توان کرد!
میلین
پوشش نورون با غلاف میلین که اسفگومیلین (عایق در برابر یون ها) دارد نشتی آن را کاهش داده و مقاومت غشا (rm) را بالا می برد ولی هرچقدر هم بالا ببرد باز هم در یک حد ثابتی می ماند چون تابع ما جذر دارد
پس باز هم جریان باید تقویت شود ولی این بار کمتر؛ این جاست که جای خالی گرههای رانویه در بحث حس میشود. گره های رانویه مکان هایی در طول نورون اند که غلاف میلین ندارند و یون ها به راحتی می توانند از عرض غشا گذرند. این احیای گسسته پتانسیل عمل را در طول غشا هدایت جهشی (saltatory condution) می گویند.
اندازه
هر چه شعاع آکسون بیشتر شود هم rm و هم ri افزایش خواهد یافت و در نگاه اول تاثیری بر ثابت طول نورون نخواهد داشت ولی کمی باید بیشتر دقت کنیم. با توجه به فرمول های دو مقاومت بالا می توانیم فرمول ثابت طول گسترش دهیم: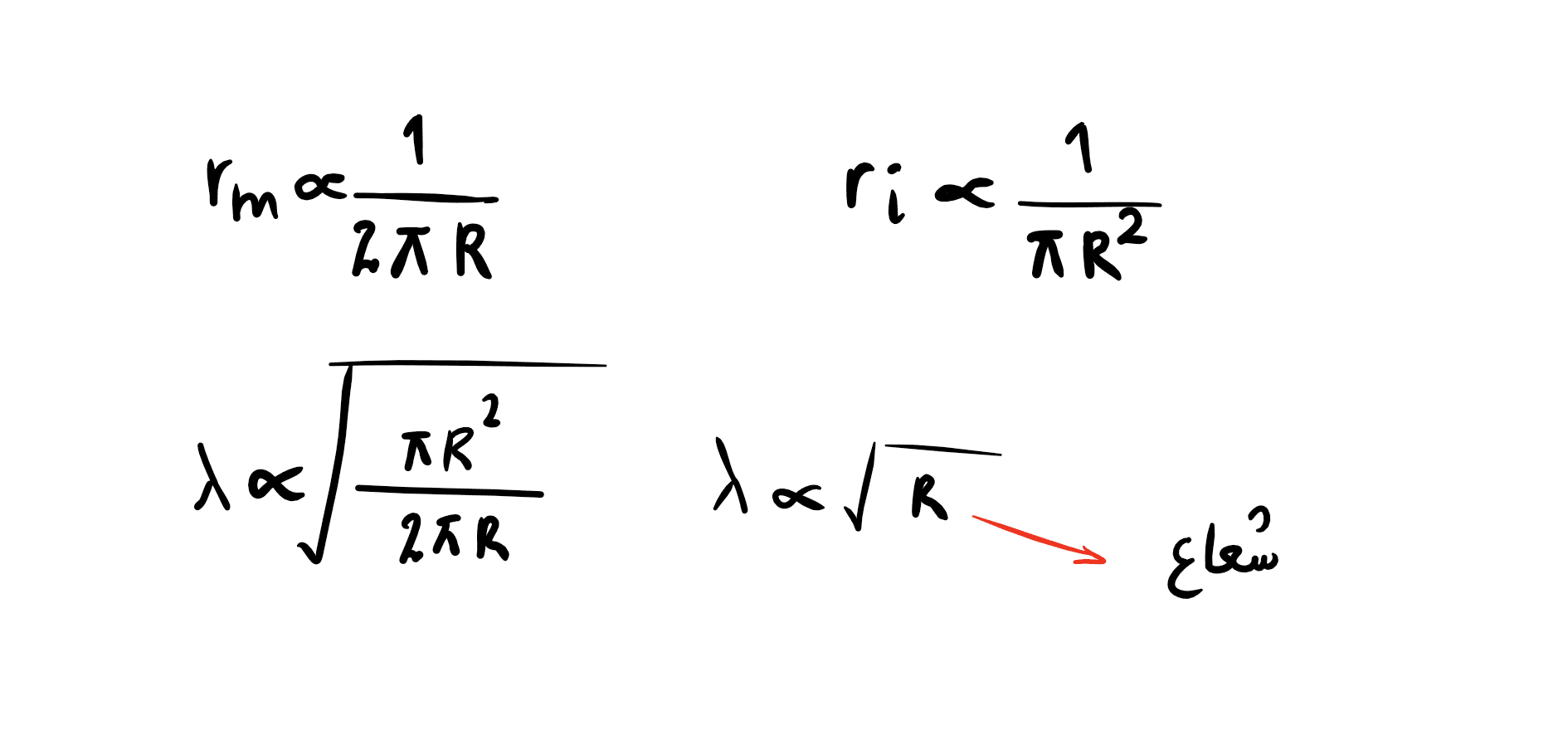
همانطور که ملاحظه میشود ثابت طول با ریشه دوم شعاع رابطه مستقیم دارد. بنابراین دو راه برای افزایش سرعت جریان وجود دارد که میلین کارایی بهتری دارد و هزینه متابولیکی آن نیز بالاست. بر همین اساس شما در بدن خود نورون هایی به شعاع چند میکرون دارید که بسیار سریع تر از نورون های چند میلی متری و بدون میلین جانوران ساده تر عمل می کنند.
- منابع:
- medical physiology Guyton & Hall
- backyardbrains
ببخشید یه سوال : کار کانل های نشتی در پتانسیل عمل چیست ؟ . آیا به کار خودوشون ادامه میدن یا نه؟ . لطفا با دلیل . سوال معلممونه.
جریان یونی از طریق کانالهای یونی بدون دریچه در عرض غشا مداوم است و به آن جریان نشتی و به کل هدایت یونی این یونها، رسانایی نشتی میگویند. در پایان پتانسیل عمل، جریان ظرفیتی مختصری به داخل سلول، دوباره غشا را رپلاریزه کرده و ولتاژ آن را به صفر برمیگرداند. (اصول علوم اعصاب کندل، صفحهی ۱۵۲)
سلام یه سوالی داشتم : ایا در زیر غلاف میلین سلول های عصبی،کانال های پروتئینی وجود دارند؟ در صورت وجود فعالیت میکنند؟
بله در زیر غلافهای میلین نیز کانالهای یونی وجود دارد ولی غلظت آنها بسیار کم است. هر چه میزان کانال کم باشد مقاومت غشا بالاتر میرود پس در مناطقی که غشای میلین وجود دارد مقاومت بسیار بالاست.
چرا نفوذ پذیری غشاءسلول عصبی نسبت به پتاسیم بیشتراز سدیم درحالت آرامش است?!
چون کانالهای همیشه باز (نشتی) پتاسیم خیلی بیشتره و کانالهای ولتاژی سدیم در حالت استراحت بسته ان؛ بنابراین نیروی کمی که بر اثر شیب الکتروشیمیایی پتاسیم ایجاد میشه میتونه جریانی رو ایجاد کنه که معادلش برای سدیم به نیروی بیشتری نیاز داره. در واقع نفوذپذیری برای تعیین نقش یون تو ایجاد پتانسیل آرامش حرف اولو میزنه (IK = gK × (Vm−EK … جی پتاسیم همون هدایت یونی یا نفوذپذیری (تعداد کانالهای یونی باز) هستش.
چراکانال های سدیم وپتاسیم جداهستند؟
اگر منظورتون این هست که چرا هر دو یون از یک نوع کانال استفاده نمیکنند؛ بخاطر تفاوت در ویژگیهای یونهاست؛ مثلاً چگالی بار یون سدیم نسبت به پتاسیم بیشتره و به همین خاطر بهتر میتونه به گروه کربونیل درون کانال بپیونده و منتقل شه. یکی دیگه از نظریهها تفاوت در قطر کانالهاست: بخاطر همون بالا بودن چگالی بار یون سدیم، مولکولهای آب بیشتری اطرافش جمع میشن (water shell) و کانالش باید قطورتر باشه. در ضمن عبور یونها از کانالهاشون نسبی عه؛ مثلا تمایل کانال پتاسیم نسبت به سدیم در مقایسه با خود پتاسیم به طور مثال یک به هزاره؛ اینطور نیست که ۱۰۰% اختصاصی باشن. با همهی اینها هنوز نظریهای نتونسته سرعت جریان ۱۰ میلیون یون بر ثانیهای از خلال کانالهای یونی رو توجیه کنه. تنوع کانالها هم خیلی بالاس، نه این که فقط دو سه تا باشه. مثلاً ژنوم ما ۹ تا متغیر ژنی برای کانال سدیمی و بیشتر از ۷۵ تا برای پتاسیمی داره.
اگر هم منظورتون از جدا بودن چرایی این اختلاف باشه؛ اصلاً همین تفاوت هست که باعث میشه پتانسیل عملی به وجود بیاد.
سلام
با توجه به توضیحاتتون پس در غشا سلول عصبی کانال نشتی سدیم نداریم؟
بعد از پتانسیل عمل آیا شیب یون های سدیم و پتاسیم مثل پتانسیل آرامشه؟ یعنی هنوز سدیم تمایل به ورود و پتاسیم تمایل به خروج داره؟؟
با تشکر از توضیحات سودمندتان
سلام ممنون از توجهتون
چرا وجود داره ولی تعدادش نسبت به کانالهای نشتی پتاسیمی بسیار کمتره
با رسیدن به قلهی پتانسیل عمل (که ۵۵+ میلیولت در نظر گرفته میشه)، شیب الکتروشیمیایی از کانالهای سدیمی وابسته به ولتاژ دیگه رو به داخل نیست و بالاتر از این حد رو به خارج میشه؛ مثل پتاسیمی.
به همین ترتیب بعد از گذراندن قله کمکم کانالهای سدیمی غیرفعال میشن و با کمی تأخیر کانالهای پتاسیمی وابسته به ولتاژ باز میشن. با خروج پتاسیم وارد فاز رپلاریزاسیون میشیم.
اما جواب سؤال اینه که تو بیشتر سلولها بعد از پتانسیل عمل یه دورهی تحریکناپذیری قطعی (عدم فعال شدن دوبارهی کانالهای وابسته به ولتاژ سدیمی) و یه دورهی تحریکناپذیری نسبی (شروع تدریجی فعالیت کانالهای وابسته به ولتاژ سدیمی در پاسخ به تحریکهای شدید و بسته شدن پتاسیمیها) رو داریم. همچنین بخاطر باز بودن کانالهای وابسته به ولتاژ پتاسیمی حتی بیشتر از حد پتانسیل آرامش غشا، پتانسیل غشا منفیتر از حالت استراحت میشه (هیپرپلاریزاسیون).
بعد از این مدت دیگه همه چی به حالت اولش (پتانسیل آرامش) برمیگرده.
سلام چرا با وجود هم بار بودن سدیم و پتاسیم ، سدیم باعث تحریک یک نورون پس سیناپس و پتاسیم باعث مهار نورون پس سیناپس میشود ؟ ممنون میشم اگر جواب بدید
با سلام
بار یون چیزی نیست که تحریک و مهار یه نورون رو تعیین بکنه. چون غلظت سدیم بیرون سلول بیشتر از درون سلوله و با رد کردن پتانسیل آستانه و باز شدن کانالهای دریچه دار نفوذپذیری غشا به اون افزایش پیدا میکنه اینطور به نظر میاد که سدیم تحریککننده اس (فاز بالاروندهی پتانسیل عمل مربوط به اونه) وگرنه پتانسیل عمل محصول تبادلات هر دو یونه.
برای درک تأثیر این دو تا یون در تحریک و مهار عصبی میتونین مثلاً مطالب مرتبط با هیپرکالمی (پتاسیم بالا) و هیپرناتریومی (سدیم بالا) رو بخونین.
دربارهی سؤال دومتون هم چیزی دربارهی این اصطلاح نشنیدم و جستجو کردمو بازم چیزی نبود. اگر میدونید معادل انگلیسی یا توضیحش رو بگید. ممنون
عذر میخوام که دوباره سوالی پرسیدم میشه لطفا درباره مرمری شدم مخ هم توضیح بدید ممنونم ☺?
سلام چرا در گره رانویه کانال نشتی وجود نداره ؟
سلام
ناحیهی خود گره (node) بسیار غنی از کانالهای دریچهدار وابسته به ولتاژ سدیم هس که «اساس هدایت جهشی در آکسونهای میلین داره». دربارهی وجود یا عدم وجود کانال نشتی در گره رانویه اطلاعی ندارم ولی در خود گره ضرورتاً کانال پتاسیمی نداریم (بیشتر کانالهای نشتی تو بقیهی جاها هم پتاسیمی عه).
وقتتون خوش معلم ما اطلاع کاملی نداد و فقط گفت که مغز بسیار شفاف و براق میشود و بقیش و ما باید تحقیق کنیم ? که من هرچی تحقیق کردم نتونستم پیدا کنم فقط استخوان مرمری آورد که اطلاعی راجب مغز نداشت
تنها مطلبی که دیدم سندرم مغز مرمری (Marble brain syndrome) بود که اونم بیماری نادر ژنتیکی با پوکی استخوان، اسیدوز کلیوی و کلیسیفیکاسیون مغزی هستش. (https://www.ncbi.nlm.nih.gov/pubmed/7358236) بهتره از متخصصان پرسوجو شه!
سلام.در کتاب ما گفته شده که نوروگلیا هادر حفظ هم ایستایی مایعد اطراف سلول های عصبی نقش دارن.چطور ممکنه که غلاف میلین هم عایق باشه و هم باعث حفظ مقدار طبیعی یون ها بشه؟
قسمت اول سؤالتون ارتباطی به قسمت دومش نداره. سلولهای نوروگلیا انواع متعددی دارن و کارای مختلفی هم انجام میدن که یکی از اونا ثابت نگه داشتن الکترولیتها مثلاً در محل سیناپس هست. به طور مثال کانال پتاسیمی دارن و این یون رو تعدیل میکنن یا به محل گرههای رانویه چسبیدن و اندازه، شکل و تراکم کانالهای یونی اون رو کنترل میکنند.
سلام وقت به خیر
میخواستم بپرسم یعنی چی که در فواصل بین گره ها هدایت پیام از نوع کاهشی است؟؟؟
باتشکر
همونطور که تو مبحث ثابت طول توضیح داده شده، با دور شدن از مجل تحریک، اختلاف پتانسیل کمتر میشه و به همین خاطر هست که به گرههای رانویه نیاز داریم تا جریان رو تقویت بکنن. به این پروسه که اختلاف پتانسیل با دور شدن از ناحیه تحریک کم میشه هدایت کاهشی میگن. برای اندازهگیریش هم با میکروالکترود زائدهی عصبی رو تحریک میکنن؛ فاصلهای از محل تحریک رو که اختلاف پتانسیل به ۳۷% مقدار اولیه برسه بهش میگن ثابت طول نورون (λ).
سلام.استاد سوالی داشتم.با توجه به اینکه در پایان پتانسیل عمل درون نورون منفی تر از حالت ارامش است در کتاب درسی یازدهم نوشته شده فعالیت بیشتر پمپ باعث می شود مجددا حالت ارامش ایجاد شود درصورتیکه پمپ سه سدیم به خارج و دو پتاسیم به داخل هدایت می کند و فعالیت بیشتر آن موجب منفی تر شدن درون باید بشود یعنی باید هیپرپلاریزاسیون را تشدید کند.آیا این عبارت کتاب درسی که فعالیت بیشتر پمپ ارامش ایجاد می کند درست است؟
با عرض سلام و ادب؛
در هنگام هایپرپلاریزاسیون، به علت منفیتر بودن پتانسیل غشا نسبت به پتانسیل نرنست یون پتاسیم، این یون به کمک کانالهای یکسو کننده به داخل سلول وارد میشود (خلاف جهت عادی) و این حرکت موجب مثبتتر شدن پتانسیل غشا و رفع هایپرپلاریزاسیون میگردد.
همچنین با توجه به جابجایی یونها حین پتانسیل عمل و بهمرور زمان، فعالیت پمپ سدیم پتاسیم به بازگشت توزیع مناسب یونها در خلال غشا کمک میکند.
سلام و درود..آیا اشتباهی در پاسخ وجود ندارد…یون سدیم هست که در پتانسیل عمل وارد سلول می شود…سپس با باز شدن کانال های ولتاژی پتاسیم پتاسیم خارج می شود تا پتانسیل عمل به حالت ارامش برگردد درست نمی گویم؟
با سلام؛
خیر اشتباهی که به آن اشاره کردید وجود ندارد. تمامی یونها نه صرفاً به دلیل شیب غلظت، بلکه به دلیل شیب الکتروشیمیایی (که شامل شیب الکتریکی + شیب غلظت) از خلال غشا به کمک کانالها عبور میکنند.
در طی Repolarization کانالهای پتاسیمی عادی باز شده و موجب خروج یون پتاسیم از درون سلول میشوند. اما هر یون در پتانسیلی مشخص از غشا، حتی در صورتی که تمام کانالهای مربوط به آن باز نیز باشد، از حرکت باز میایستد. این پتانسیل به «پتانسیل نرنست آن یون» معروف است. لذا یون K تا پیش از Hyperpolarization به دلیل مثبتتر بودن پتانسیل غشا نسبت به پتانسیل نرنست پتاسیم به خارج از سلول رفته و موجب منفیتر شدن پتانسیل غشا میگردد. اما با شروع فرایند Hyperpolarization و منفیتر شدن پتانسیل غشا نسبت به نرنست پتاسیم، این یون به کمک کانالهای یکسو کننده یا KIR یا Inward Rectifier Potassium Channels، «وارد» سلول شده و موجب مثبت شدن پتانسیل غشا و کمک به خاتمهی فرایند Hyperpolarization میگردد.
و اگر سدیم وارد می شود چرا بر خلاف شیب…مگر در بیرون بیشتر نیست؟ پس باید در جهت شیب باشد…
سلام یه سوالی داشتم
با توجه به مراحل جداسازی سدیمی و پتاسیمی نحوه به دست امدن جریان سدیم را شرح دهید
سلام
منظور از منفی کردن پتانسیل پردازش شبکه ای پس سیناپسی چیست.
ببخشید میتونیم بگیم کانال های نشتی سدیم … میتوانند سدیم خارج کنند مثلا در هنگام پتانسیل عمل که غلظت سدیم داخل بیشتره اینا خارج میکنن سدیم رو
جریان یونی کانالهای نشتی همیشه برقراره
ولی غلظت سدیم داخل سلول هیچ وقت بیشتر از خارج سلول نمیشه؛ حتی تو پتانسیل عمل.
تا ۵۵+ شیب شیمیایی رو به داخل و شیب الکتریکی رو به بیرون عه ولی شیب شیمیایی بزرگتر از الکتریکی عه و بنابراین سدیم داخل سلول میشه
ولی بالاتر از ۵۵ شیب الکتریکی به شیمیایی غلبه کرده و سدیم را از سلول خارج میکند.
سلام
راجع به ریپلاریزاسون ، دپلاریزاسیون و هایپرپلاریزاسیون میشه توضیح بدید به چه معنی هستند؟
و راجع به refractory period
با تشکر
کسی میتونه انواع کانال های سلول را بطور کامل برام بگه
خیلی از کانالهای یونی رو در غشای سلول میشه تو یکی از این چهار تا دسته قرار داد:
۱. کانالهای ولتاژی (voltage gated)؛ در پاسخ به تغییر پتانسیل غشا باز و بسته میشوند. کانالهای دریچهدار سدیمی و پتاسیمی که تو پتانسیل عمل نقش دارند جزو این دسته هستن.
۲. کانالهای لیگاندی (ligand gated)؛ در پاسخ به مواد شیمیایی باز و بسته میشوند. مثلاً در پاسخ به استیلکولین و …
۳. کانالهای مکانیکی (mechanically gated)؛ فشار و کشش باعث باز و بسته شدنشون میشه؛ مثلاً کانالهای کلسیمی که انقباض میوتونیک رو باعث میشن.
۴. کانالهای نشتی یا استراحت (resting)؛ در حالت آرامش سلول با نشت یونها از این کانالها، کمک مهمی به ایجاد پتانسیل آرامش میشود. مثل کانالهای نشتی پتاسیمی و سدیمی
قطر کانال های سدیم پتاسیمی چه تغییری میکنند زمانی که وارد محیط ابی میشه
با سلام سوال این هست که: تعداد دریچه های کانال های یونی در واحد سطح چقدر است؟
همچنین تعداد کانالهای یونی در واحد سطح چقدر است؟؟؟
با سلام
تعداد دریچههای یونی برای هر کانالی یکی یا دوتاست. در ضمن دریچهها فقط مدل هستن و در واقعیت دریچهای وجود نداره. اما سؤال دومتون:
تعداد کانالهای یونی در واحد سطح غشا رو نمیشه مستقیم شمرد و اندازه گرفت! ولی با در دست داشتن مقاومت یک کانال و مقاومت سطحی از غشا، میتوان تعداد کانالهای “باز” را در آن سطح تخمین زد. ما یک مفهوم داریم دربارهی نورون، به نام “ثابت زمان”؛ ثابت زمان به مدت زمانی میگن که یه جریان بدون تحریک دوباره در گرهها میتونه زنده بمونه. حالا چه ربطی به سؤال شما داره؟ خب فرمول اینه:
ثابت زمان= ظرفیت غشا * مقاومت غشا
ثابت زمان رو میشه تو آزمایشگاه اندازه گرفت و چیزی بین ۱۰ میلیثانیه تا ۱ ثانیهاس. ظرفیت غشا هم واس همهی غشاهای زیستی ۱ میکروفاراد بر سانتیمتر مربع هست.
مثلاً برای پارامسی رو اندازه گرفتن و مقاومتش ۶۰,۰۰۰ اهم بر سانتیمتر مربع اومده. با این حساب اگه مقاومت یک کانال رو ۱۰ به توان ۱۲ در نظر بگیریم، در هر سانتیمتر مربع از غشای پارامسی، چیزی در حدود ۱۶ میلیون کانال بونی وجود داره.
پ ن: ممکنه مقاومت یک کانال رو جاهای دیگه به صورت متفاوتی گزارش کرده باشن.
سلام خسته نباشید. یک سوال داشتم:
چه چیزی باعث میشه که فقط سدیم ها از کانال دریچه دار سدیمی عبور کنند و فقط پتاسیم ها از کانال دریچه دار پتاسیمی؟ آیا این امکان وجود دارد که از کانال های یکدیگر عبور کنند؟ چه چیزی باعث ایجاد این نظم و مانع عبور یون دیگر می شود؟ ممنون میشم سریع تر پاسخ بدید دارم از کنجکاوی می میرم .
با سلام
بخاطر تفاوت در ویژگیهای یونهاست؛ مثلاً چگالی بار یون سدیم نسبت به پتاسیم بیشتره و به همین خاطر بهتر میتونه به گروه کربونیل درون کانال بپیونده و منتقل شه. یکی دیگه از نظریهها تفاوت در قطر کانالهاست: بخاطر همون بالا بودن چگالی بار یون سدیم، مولکولهای آب بیشتری اطرافش جمع میشن (water shell) و کانالش باید قطورتر باشه. در ضمن عبور یونها از کانالهاشون نسبی عه؛ مثلا تمایل کانال پتاسیم نسبت به سدیم در مقایسه با خود پتاسیم به طور مثال یک به هزاره؛ اینطور نیست که ۱۰۰% اختصاصی باشن. با همهی اینها هنوز نظریهای نتونسته سرعت جریان ۱۰ میلیون یون بر ثانیهای از خلال کانالهای یونی رو توجیه کنه.
سلام
می دونیم که فعالیت پمپ سدیم پتاسیم از نوع انتقال فعال هستش ، یعنی سدیم رو از داخل که غلظت کمتری داره به بیرون و پتاسیم رو که در بیرون غلظت کمتری داره به داخل سلول هدایت می کنه و چون این انتقال در خلاف جهت شیب غلظت هست مجبوره ATP مصرف کنه
حالا سوالم اینه که بعد از پایان پتانسیل عمل که جای یونها تقریبا عوض شده و در داخل بیشتر سدیم داریم و در خارج بیشتر پتاسیم، دوباره پمپ سدیم-پتاسیم فعال میشه و سدیم رو به خارج و پتاسیم رو به داخل هدایت می کنه
حالا توی این مرحله هم باز کار این پمپ از نوع انتقال فعاله؟
چون وقتی مثلا داخل سلول سدیم زیاد شده و پمپ باید سدیم رو به خارج هدایت کنه ، دیگه در خلاف شیب غلظت نیست و در واقع میشه در جهت شیب غلظت. ولی میدونیم که انتقال فعال در خلاف شیب غلظت هست
چون توی بعضی از سایتها و منابع معتبر دیدم و خوندم که نوشتن بعد از پایان پتانسیل عمل ، پمپ سدیم-پتاسیم با مصرف ATP فعال میشه (انتقال فعال) و جای یونهای سدیم و پتاسیم رو به حالت اولیه خودش برمیگردونه
آیا امکانش هست پمپ سدیم -پتاسیم در حالت آرامش از طریق انتقال فعال کار کنه(ینی خلاف شیب غلظت) و بعد از پتانسیل عمل در جهت شیب غلظت (ینی انتشار)؟
اگر انتشار هست پس دیگه چرا انرژی مصرف میکنه؟و اگه انتقال فعال هست پس شیب غلظت به کدوم سمت هست؟
ببخشید که سوالم طولانی شد
سلام
قبل از جواب باید اشاره کنم که توی سؤال شما دو تا اشکال وجود داره:
۱. در داخل همواره پتاسیم بیشتر است؛ این فقط در ابر یونی اطراف غشاست که جای یونها عوض میشود
۲. پمپ سدیم/پتاسیم همیشه فعاله، نه فقط بعد از پایان پتانسیل عمل
بعد از رپلاریزاسیون که کانالهای دریچهدار پتاسیمی باز میشن و پتاسیم از سلول خارج میشه، سلول هایپرپلاریزه میشه؛ یعنی پتانسیل غشا از پتانسیل نرنست پتاسیم هم کمتر میشه. حالا چه اتفاقاتی باعث میشه این فرآیند خاتمه پیدا کنه و به حالت آرامش برگرده؟
در جواب باید گفت که یک عامل وجود نداره. هم کانالهای یکسوکنندهی پتاسیمی (Inward Rectifier Potassium Channels) فعال میشن و پتاسیم رو وارد سلول میکنن، هم کانالهای نشتی دوباره کارشونو از سر میگیرن و پتاسیم رو خارج و سدیم رو وارد میکنن، هم یون کلر نقش داره و در این بین، پمپ سدیم/پتاسیم هم به موندن در پتانسیل استراحت کمک میکنه. کار پروتئینها اختصاصیه و حداقل در این مورد نمیتونه تغییر کنه؛ پس همواره انتقال فعال رو انجام میده. موفق باشین.
سلام وقت تون بخیر سئوالی ذهنم رو درگیر کرده و سایت ها و منابع زیادی رو به دنبال پاسخ گشتم اما دقیقا متوجه نشدم اینکه چطور جابجایی یون ها یعنی هجوم یون سدیم به داخل و یون پتاسیم به خارج توسط کانال های نشتی باعث یک جریان الکتریکی میشه و دقیقا چطور این جابجایی باعث تحریک مکان بعدی (یا نقطه بعدی در همان سلول) میشه؟
با تشکر.
سلام
ورود سدیم به داخل سلول غشا رو دپلاریزه میکنه؛ یعنی برخلاف بقیه جاها این مکان درونش مثبت و بیرونش منفی ئه. حالا چطور تبدیل به جریان میشه؟ با فیدبک مثبت. در محلی از غشا که دپلاریزه شده، کانالهای سدیمی اطراف اون هم به آستانهی تحریک میرسن و جریان در هر دو طرف محل تحریک به سمت انتهاها پیش میره
دقیقا همین نکته برای من سئواله که چطور با دپلاریزه شدن محلی از غشا کانال های سدیمی اطرافش هم به آستانه تحریک می رسند؟ مخصوصا زمانی که در مورد گره های رانویه که کانال های سدیمی از هم فاصله دارند صحبت می کنیم.
کانالهای سدیمی وابسته به ولتاژن (voltage gated). یعنی وقتی ولتاژ دو سوی غشا تغییر میکنه باز و بسته میشن. شما “فرض کنید” دریچهی کانال سدیمی بار مثبت داره. وقتی غشا در پتانسیل آرامشه، طرف داخل منفیه؛ این یعنی دریچه و یونهای طرف داخل همدیگه رو جذب میکنن و کانال بسته میمونه.
وقتی تعدادی از کانالها “بر اثر تحریک خارجی” باز بشن دپلاریزاسیون اتفاق میوفته و طرف داخل مثبت میشه. خوب الآن مثبتِ داخل، دریچه مثبت رو دفع میکنه و بقیهی کانالا هم باز میشن و به همین شیوه (فیدبک مثبت) تا یه حدی بقیه کانالها هم باز میشن و جریان ایجاد میشه. برای اینکه بهتر جا بیفته به این شکل دقت کنین (https://goo.gl/zk7GxE)
در مورد گرههای رانویه هم باید بگم که تعداد کانالها در بین دو گره کم هست نه اینکه نباشه و به شیوهای که گفتیم تحریک جلو میره. اگه کامل خونده باشین مقاله رو، ثابت طول رو هم خوندین و پس از طی اون مسافت جریان به گره میرسه و با تعداد زیاد کانالهای سدیمی دوباره تقویت میشه. این شکل کمکتون میکنه (https://goo.gl/4q2wSY)
متشکرم، تا حدودی متوجه شدم و در کل به این شکل هدایت پیام عصبی رو استدلال کردم که با تحریک خارجی مقداری یون های سدیم با بار مثبت به کانال نفوذ می کنند و داخل مثبت تر و اختلاف پتانسیل الکتریکی کاهش می یابد و به کمک همین سدیم ها کانال ها باز شده و انبوهی از یون های سدیم به داخل نفوذ می کنند برای هدایت هم مقداری از سدیم ها داخل آکسون به نقطه بعدی می روند و کانال ها را باز می کنند و دوباره پتانسیل عمل اتفاق می افتد…
سلام.خسته نباشید.
بخشید وقتی که پتانسیل غشا از ۷۰- به ۳۰+ میرسد کاهش میابد یا افزایش.
و کانال های نشتی،تقسیم میشود به کانال های نشتی پتاسیمی و کانال های نشتی سدیمی؟ در برخی کتاب ها به نشت سدیم از کانال های نشتی سدیم-پتاسیمی اشاره دارد، کدوم درسته؟
سلام
“اختلاف پتانسیل بین دو سوی غشا” در دپلاریزاسیون ابتدا کاهش و سپس افزایش پیدا میکنه
تا جایی که تو منابع نوشته شده کانالهای نشتی سدیم و پتاسیم جدا هستند. درسته عبور یون از این کانال مطلقاً اختصاصی نیست و تا حد بالایی اختصاصی کار میکنن
کسی میتونه جواب بده؟
با سلام. میخواستم بدونم وقتی یک بافت تخریب یا آسیب میبیند سلول های سالم از کجا میفهمند چقدر باید میتوز کنند و یا کدام سلول ها مسول این کارند؟ اگر درست فکر کنیم همه سلول های سالم نزدیک به قسمت آسیب دیده که تقسیم نمیشن. تازه! چند بار باید تقسیم شن رو چه مکانیسمی کنترل میکنه؟ برام خیلی سواله.
ممنون میشم جواب بدین.
باتشکر از سایت مفید و در عین حال با طراحی عالی تر نسبت به بقیه سایت ها.
سلام
وقتی بافتی در بدن آسیب میبیند، معمولاً پای سیستم ایمنی به میان کشیده میشود. اجزای سیستم ایمنی پس از سرکوب عفونت احتمالی، یا شیء خارجی (و سایر نقشهای ایمنی) به کار ترمیم میپردازند. در این مسیر، به طور مثال، ماکروفاژها از مسیر دوم فعال شده و با “ترشح انواع سایتوکینهای ترمیمی” مثل TGF B، به سایر سلولها پیام تکثیر و تقسیم میفرستند. در کل، این وظیفهی سیگنالینگ سلولی، سایتوکینها، هورمونها و… است که پیام رشد، ترمیم، مهار رشد و غیره را به سلولها برسانند. یا مثلاً این VEGF است که باعث تحریک رگزایی میشود.
در مورد تشکیل اسکار (بافت زخم) هم سلولهای فیبروبلاست و فیبر اصلی آنها، کلاژن نقش دارند. در ضمن سلولهای بنیادی در بسیاری از بافتها در پاسخ به این سیگنالها تکثیر شده و جای سلولهای از دست رفته را میگیرند؛ این کار در بافتهایی که دائم در حال فرسایش اند، مثل مجرای گوارشی، فیزیولوژیک است.
در مورد چند بار تقسیم شدن سلولها هم باید چرخهی سلولی رو مطالعه کنین https://goo.gl/edzKLT
یه جا خوندم میگفت نورووگلیا پتانسیل ارامش داره یعنی اونم اختلاف پتانسیل دو سوی غشاش ۷۰-هست؟؟
اگر کانال های دریچه دار پتاسیمی بیش از حدباز بمانند اختلاف پتانسیل چه تغییری میکنه؟
هر کانالی که بیشتر باز بمونه، نفوذپذیری اون یون رو افزایش میده، و هر یونی نفوذپذیری بیشتری داشته باشه پتانسیل غشا رو به پتانسیل نرنست خودش نزدیکتر میکنه؛ یعنی اگه پتاسیمی بیشتر باز بمونه، سلول هایپرپلاریزه میشه
سلام فرق پمپ، پروتئین ناقل و پروتئین حامل چیه؟ هر کانال نشتی که تو غشا هست مخصوص یک یون اختصاصی است و بس؟