عناصر قابل جابهجایی (Transposable elements) یا ترانسپوزون ها که عنوان “jumping genes” را نیز برای آنها به کار میبرند، توالیهایی از DNA هستند که مکان خود را روی ژنوم عوض میکنند. این عناصر برای اولین بار بیش از ۵۰ سال پیش، توسط متخصص ژنتیک، باربارا مککلینتوک از آزمایشگاه Cold Spring Harbor در نیویورک شناسایی شدند. زیستشناسان ابتدا با دیدهی تردید به این کشف نگریستند ولی در طی دهههای بعدی مشخص شد که این عناصر نهتنها خیلی متحرک اند، بلکه تقریباً در همهی ارگانیسمها (هم پروکاریوتها و هم یوکاریوتها) وجود دارند و کم هم نیستند. برای مثال، عناصر قابل جابهجایی حدوداً ۵۰ درصد ژنوم انسان و تا ۹۰ درصد ژنوم ذرت را تشکیل میدهند.
انواع ترانسپوزونها
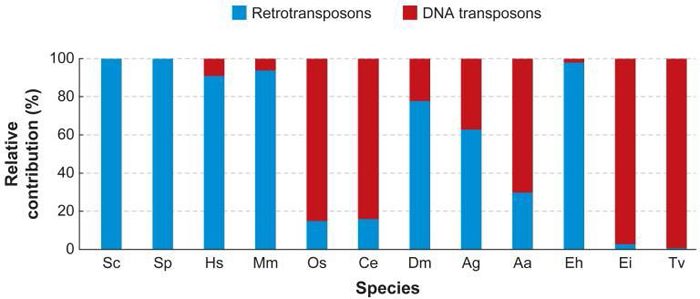
امروزه دانشمندان به این نتیجه رسیدهاند که انواع مختلفی از TE وجود دارد؛ راههای طبقهبندی هم از لحاظ تنوع دست کمی از خود ترانسپوزونها ندارد. یکی از تقسیمبندیهای مرسوم بر اساس این است که ترانسپوزون برای جابهجایی به رونویسی معکوس (DNA ساختن از RNA) نیاز دارد یا نه. عناصر سابق را با نام رتروترانسپوزونها یا TEهای کلاس یک میشناسیم؛ در حالی که بقیه با نام DNA ترانسپوزون یا TEهای کلاس دو شناخته میشوند. سیستم Ac/Ds که مککلینتوک کشف کرده بود در زمرهی کلاس دو قرار میگیرد. کلاسهای متفاوتی از عناصر قابل جابهجایی (TEs) در ژنوم انواع ارگانیسمهای یوکاریوتی یافت میشود (شکل ۱).
DNA ترانسپوزونها
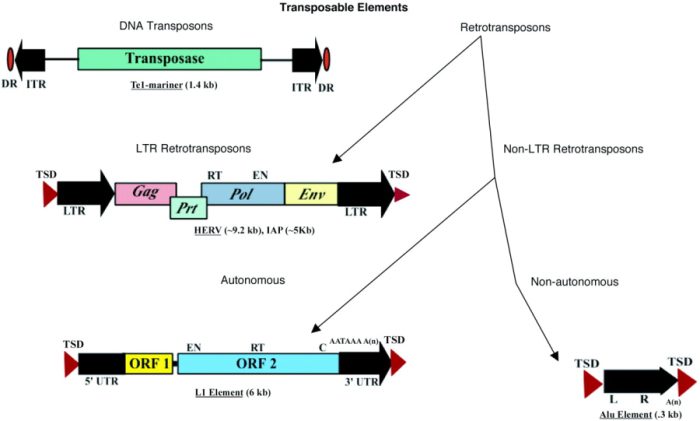
کلاس دو عناصر قابل جابهجایی پروتئین ترانسپوزاز را کد میکنند که برای برش و تعبیه نیاز است (شکل ۲). برخی از این عناصر نیز پروتئینهای دیگری را کد میکنند. به یاد داشته باشید که DNA ترانسپوزونها هرگز از واسطههای RNA استفاده نمیکنند؛ آنها همیشه به خود متکی اند. برش و تعبیهی آنها در ژنوم از طریق مکانیسم “برش و چسباندن” انجام میگیرد.
عناصر قابل جابهجایی کلاس دو با حضور تکرارهای انتهایی معکوس، به طول ۹ تا ۴۰ جفت باز در دو طرف مشخص میشوند (شکل ۳). همانطور که از اسم برمیآید و شکل ۳ نیز نشان میدهد، تکرارهای انتهایی معکوس، مکملهای معکوس همدیگر هستند؛ برای مثال، مکمل ACGCTA (تکرار معکوس در طرف راست TE در شکل) TGCGAT است. یکی از نقشهای این توالیها، شناسایی توسط ترانسپوزازهاست.
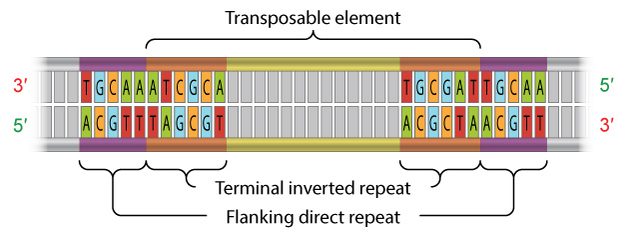
علاوه بر این، همهی ترانسپوزنهای کلاس یک و دو حاوی تکرارهای مستقیم جانبی میباشند (شکل ۳). تکرارهای مستقیم جانبی جزء مسلم ترانسپوزونها نیستند؛ به جای آن، نقش این توالیها در تعبیهی ترانسپوزونهاست. فراتر از این، پس از جدا شدن ترانسپوزون، این توالیها نقش “ردپا” را دارند. برخی اوقات، این ردپاها بیان ژن را تغییر میدهند (بیان ژنی را که ترک کردهاند)؛ حتی بعد از حرکت کردن ترانسپوزن از مکان اولیهی خودش.
کمتر از دو درصد ژنوم انسان ترانسپوزن کلاس دو در نظر گرفته میشود. این به آن معناست که عمده گروههای ترانسپوزونی را رتروترانسپوزونها تشکیل میدهند.
رتروترانسپوزونها
برخلاف عناصر کلاس دو، عناصر کلاس یک که با نام رتروترانسپوزونها هم شناخته میشوند، جای خالی واسطههای RNA را پر میکنند. به عبارتی دیگر، ترانسپوزونهای کلاس یک ترانسپوزاز کد نمیکنند؛ به جای آن، رونوشتهایی از RNA تولید میکنند که سپس به کمک آنزیمهای رونویسی معکوس (RTase) توالی RNA را به DNA تبدیل میکند که قرار است در مکان هدف قرار گیرد.
دو گروه اصلی ترانسپوزون کلاس یک وجود دارد: رتروترانسپوزونهای LTR، که با تکرارهای انتهایی طولانی (LTRs) در دو سمت مشخص میشوند. و رتروترانسپوزونهای non-LTR که توالی مذکور را ندارند. هم ژن (LINE1 (L1، و هم ژنهای Alu نمایندهی خانوادههای ترانسپوزونهای non-LTR میباشد. عناصر L1 به طور میانگین طولی به اندازهی شش کیلوباز دارند. برخلاف آن، عناصر Alu فقط چند صد نوکلئوتید دارند و این کوتاهی از آنها عناصر قابل جابهجایی درهمآمیختهی کوتاه (SINE) میسازد.
ژن Alu به طور ویژهای فراوان است؛ در نخستیها به وجود آمده و در بازهی زمانی به نسبت کوتاهی تقریباً یک میلیون رونوشت در هر سلول انسانی ایجاد کرده است. L1 هم در انسان ژن فراوانی میباشد؛ با این که تعداد L1 به Alu نمیرسد ولی به خاطر اندازهی بزرگترش، ۱۵ تا ۱۷ درصد ژنوم را انسان را به خود اختصاص داده است. در انسانها، این ترانسپوزونهای non-LTR، تنها کلاسهای فعال از ترانسپوزونها هستند؛ رتروترانسپوزونهای LTR و DNA ترانسپوزونها فقط آثاری از ژنومیک باستانی اند و قابلیت حرکت در آنها وجود ندارد.
ترانسپوزونهای خودمختار و غیرخودمختار
هم ترانسپوزونهای کلاس یک و هم کلاس دو میتوانند خودمختار (autonomous) یا غیرخودمختار (nonautonomous) باشند. ترانسپوزونهای خودمختار میتوانند به خودی خود حرکت کنند؛ در حالی که عناصر غیرخودمختار برای حرکت به ترانسپوزونهای دیگر وابسته اند. این به خاطر عدم حضور ژن ترانسپوزاز یا رونویسیکنندهی معکوس میباشد که برای تغییر موقعیت آنها الزامی اند؛ بنابراین آنها باید این پروتئینها را قرض بگیرند. برای مثال، عناصر Ac خودمختارند، چون میتوانند خودشان حرکت کنند؛ بالعکس، عناصر Ds غیرخودمختارند؛ چون برای تغییر موقعیت به حضور Ac نیاز دارند.
ژنهای معلق چه کاری انجام میدهند (البته به جز معلق بودن)؟!
این واقعیت که تقریباً نیمی از ژنوم ما را ترانسپوزونها تشکیل میدهند و قسمت اعظم آنها نیز رتروترانسپوزونهای L1 و Alu است، سؤال مهمی را ایجاد میکند: همهی این ژنهای معلق، به جز معلق بودن، چه کاری انجام میدهند؟ جواب این سؤال بیشتر به مکانی بستگی دارد که ترانسپوزون در آن جا قرار میگیرد. قرار گرفتن وسط یک ژن، میتواند به یک جهش منجر شود؛ همانطور که کشف شد، قرارگیری L1 در ژن فاکتور VIII انعقادی موجب ایجاد هموفیلی میگردد.
به همین شیوه، چند سال بعد، محققان رد ژن L1 را در بین ژنهای APC در سلولهای کولونی سرطانی گرفتند؛ در حالی که در سلولهای سالم همان افراد چنین چیزی مشاهده نمیشد. این مشاهدات تأیید میکنند که L1 در سلولهای پیکری پستانداران تغییر موقعیت میدهد، و اینکه این عنصر شاید نقشی در ایجاد و رشد بیماریها داشته باشد.
سرکوب ترانسپوزونها
بر خلاف L1، به نظر میرسد بیشتر ترانسپوزونها خاموش باشند. به عبارتی دیگر، این عناصر نه اثرات فنوتیپی دارند و نه در ژنوم حرکت میکنند. حداقل اجماع علمی بر این است. برخی از ترانسپوزونها خاموش شدهاند؛ چون جهشهایی دارند که توانایی انتقال بین کروموزومها را از آنها گرفته است. بقیه کاملاً دستنخورده و دارای توانایی حرکت اند، ولی به واسطهی مکانیسمهای دفاعی اپیژنتیک نظیر متیلاسیون DNA، ریمدلینگ کروماتین و میکروRNAها غیرفعال نگه داشته شدهاند. مثلاً در ریمدلینگ کروماتین، تغییرات شیمیایی در پروتئینهای کروماتین موجب فشرده شدن آن در برخی نواحی ژنوم میشود که ترانسپوزونها وجود دارند. به همین دلیل آنزیمهای رونویسیکننده به راحتی نمیتوانند به آنها دسترسی پیدا کنند.
مثال دیگر از خاموش کردن ترانسپوزون، گیاهانی در جنس آرابیدوپسیس است. محققان در این گیاهان، بیش از ۲۰ توالی جهشدهندهی ترانسپوزون یافتهاند (نوعی ترانسپوزون که در ذرت شناسایی شده است). در گیاهان وحشی، این توالیها متیله، یا خاموش شدهاند. با این حال، گیاهانی هم وجود دارند که در آنزیمهای مسئول متیلاسیون نقص دارند. ترانسپوزونها در این گیاهان فنوتیپهایی را سبب شدهاند.
بر اساس مطالعات چنینی، دانشمندان میدانند که برخی ترانسپوزونها به صورت اپیژنتیکی خاموش میشوند؛ در سالهای اخیر، محققان توجه خود را به نقش خود ترانسپوزونها در خاموش شدنشان متمرکز نمودهاند. جالب است بدانید که این خود باربارا مککلینتوک بود که این نقش تنظیمی احتمالی را معرفی کرد. ولی دههها زمان برد تا شواهد کافی برای آن جمعآوری شود.
ترانسپوزونها میتوانند siRNAهایی کد کنند که خاموش شدن خودشان را تعدیل میکند
چون حرکت ترانسپوزون میتواند تخریبکننده باشد، جای تعجب نیست که بیشتر توالیهای ترانسپوزون در ژنوم انسان خاموش شده است؛ این کار، علیرغم وجود ترانسپوزونهای فراوان به ثبات نسبی ژنوم کمک میکند. در واقع، محققان بر این باورند که از ۱۷ درصد کل ژنوم انسان که توسط توالیهای مرتبط با L1 کدگذاری میشود، تقریباً فقط ۱۰۰ عنصر فعال L1 باقی مانده است. تحقیقات جدید راههای بیشتری را نسبت به تغییرات اپیژنتیک، به منظور جلوگیری از جهش ترانسپوزونها پیشنهاد میکند.
برای نمونه، در سلولهای انسانی، RNAهای تداخلگر کوچک (siRNAs)، که با مخفف RNAi هم شناخته میشوند، میتوانند از ترانسپوزیشن جلوگیری کنند. siRNA مکانیسم ذاتی در یوکاریوتهاست که اغلب برای تنظیم بیان ژن استفاده میشود. جالب توجه این است که siRNAهایی که در فعالیت L1 تداخل میکنند، از منطقهی ترجمهنشدهی ۵′ (۵′ UTR) مربوط به L1 LTR منتقل میشود. مخصوصاً، ۵′ UTR مربوط به پروموتور L1 پروموتور سنسی را کد میکند که از ژنهای L1 رونویسی خواهد کرد؛ همچنین پروموتور آنتیسنسی هم به وجود میآید که RNA آنتیسنسی را خواهد ساخت. یانگ و کازازیان (۲۰۰۶) نشان دادند که این اتفاق به توالیهای همولوگی میانجامد که میتوانند هیبریده شده و RNA دو رشتهای را بسازند که پیشساز RNAi باشد. علاوه بر اینها، وقتی محققان مکانیسمهای خاموشسازی siRNA اندوژن را مهار کردند، رونویسی از L1 افزایش یافت؛ این مشاهدات پیشنهاد میکند که رونویسی از L1is توسط siRNA مهار میشود.
ترانسپوزونها همیشه هم تخریبکننده نیستند
همهی جهشهای ترانسپوزونی عواقب تخریبگر ندارد. در واقع، ترانسپوزونها میتوانند تکامل ژنومها را با تسهیل ترانسلوکاسیون توالیهای ژنومیک، پخش کردن اگزونها و ترمیم شکستگیهای دورشتهای بدست بگیرند. ترانسپوزیشن همچنین میتواند مناطق تنظیمی ژن و فنوتیپها را تغییر دهد. برای نمونه، در ماهی مداکا، DNA ترانسپوزون Tol2 مستقیماً با پیگمانتاسیون ارتباط دارد. یکی از این نوع ماهیها به طور ذاتی الگوهای پیگانتاسیون متنوعی دارد. در ماهیهایی که این ترانسپوزون بدون آسیب زدن به ژنهای اطراف حذف شده است، زالی دیده میشود. ولی وقتی Tol2 تر و تمیز حذف نشود، نتیجه میشود طیفی وسیع از الگوهای پیگمانتاسیون ارثی.
این واقعیت که عناصر قابل جابهجایی همیشه کاملاً دقیق برش نمیخورند و میتوانند توالیهایی را هم با خود همراه کنند، به پدیدهای انجامیده که دانشمندان به آن آمیختگی اگزون میگویند. آمیختگی اگزون معمولاً با ترانسپوزیشن به همجوار شدن دو اگزون غیرمرتبط به هم منتج میشود که به طور بالقوه محصولات ژنی جدیدی به وجود میآورد.
قابلیت ترانسپوزونها در افزایش تنوع ژنتیکی، به همراه توانایی ژنوم در مهار فعالیت آنها، به تعادلی میانجامد که این عناصر را به یکی از مهمترین قسمتهای تکامل تنظیم ژن در همهی ارگانیسمهای حاوی این توالیها بدل کرده است.