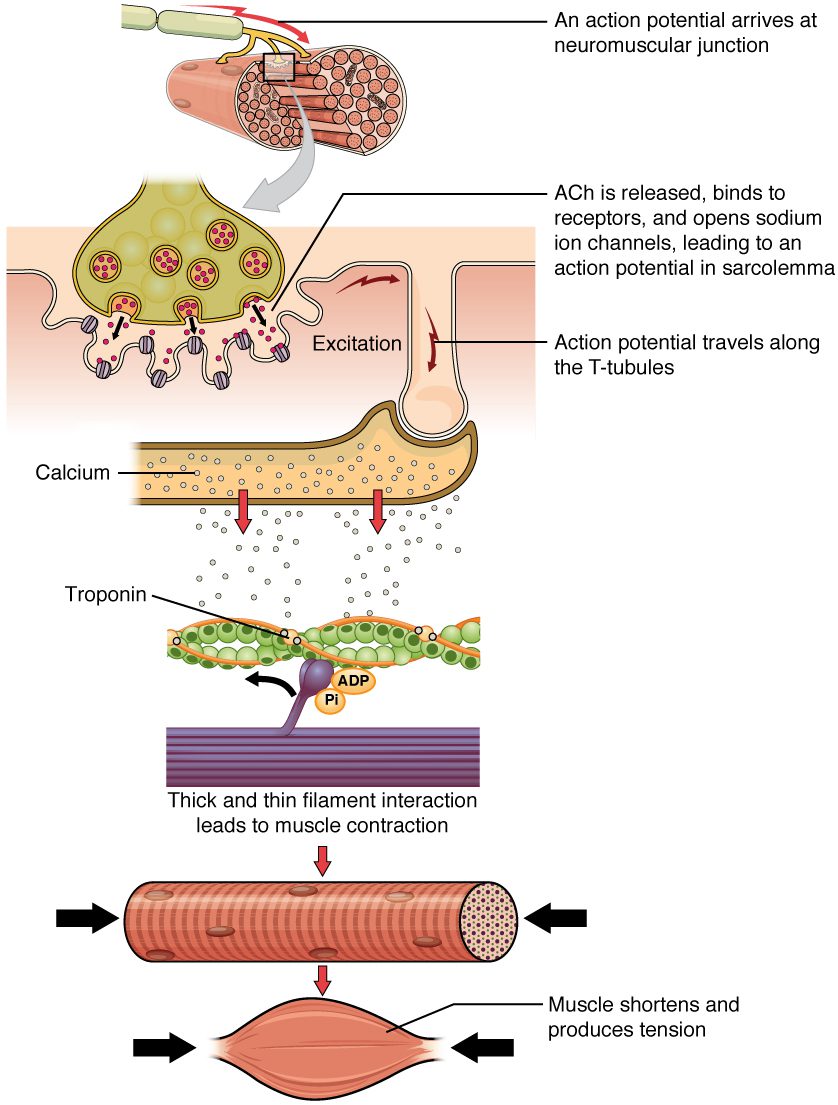
مکانیسم انقباض ماهیچه مخطط مجموعه رویدادهایی است که همگی از یک سیگنال آغاز میگردند. این سیگنال، نوروترنسمیتر استیل کولین است و از نورون حرکتی عصبدهی کننده آن فیبر آزاد میشود. در این حالت همزمان با ورود یونهای مثبت سدیم، غشای فیبر به صورت موضعی دپلاریزه می شود. درنتیجه ایجاد پتانسیل عمل تحریک و این پتانسیل در سرتاسر غشا از جمله توبولهای T منتشر میگردد. این موضوع باعث رها شدن یونهای کلسیم از شبکه سارکوپلاسمی میشود. یونهای کلسیم سپس فرایند انقباض را که از ATP انرژی میگیرد، آغاز میکنند. تا زمانی که یونهای کلسیم در سارکوپلاسم وجود داشته باشند تا به تروپونین متصل شوند و جایگاههای اتصال اکتین باز باقی بمانند، و تا زمانی که ATP موردنیاز برای چرخه پلهای عرضی و کشیده شدن رشتههای اکتین توسط میوزین فراهم باشد، فیبرهای عضلانی به کوتاه شدن در محدوده آناتومیک ادامه خواهند داد.
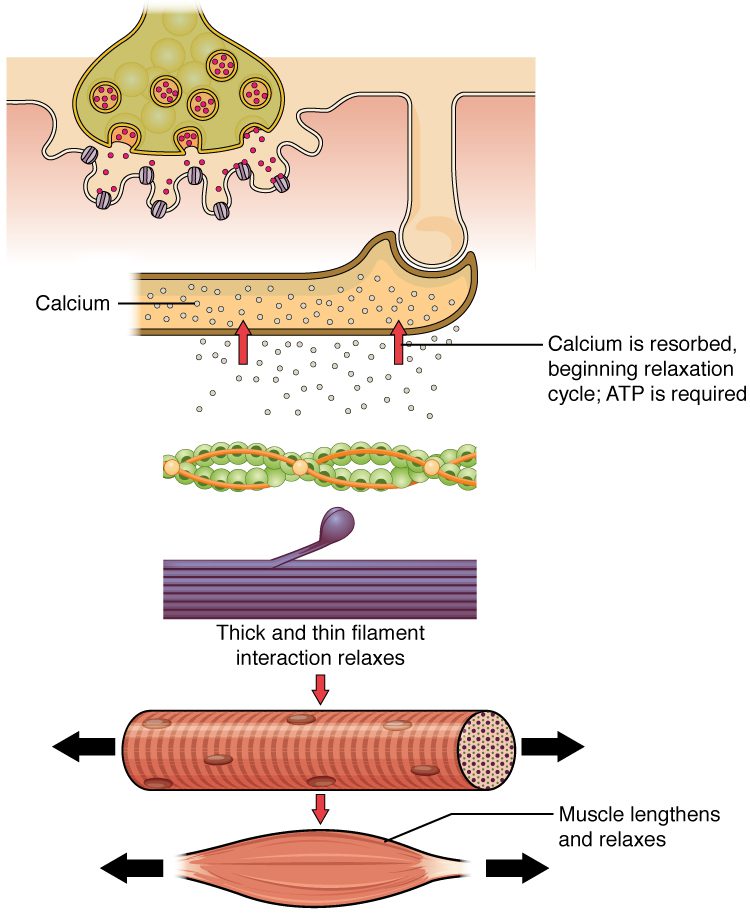
انقباض عضلات با توقف سیگنال آزاد شده از انتهای نورونهای حرکتی متوقف میشود. توقف سیگنال منجر به رپلاریزه شدن سارکولما و توبولهای T و بسته شدن کانالهای کلسیمی وابسته به ولتاژ در شبکه سارکوپلاسمی میگردد. یونهای کلسیم سپس مجدددا به داخل شبکه سارکوپلاسمی پمپ میشوند. درنتیجه تروپومیوزین مجددا جایگاههای اتصال موجود در اکتین را میپوشاند. توقف انقباض میتواند در صورت اتمام ATP و خستگی نیز رخ دهد.
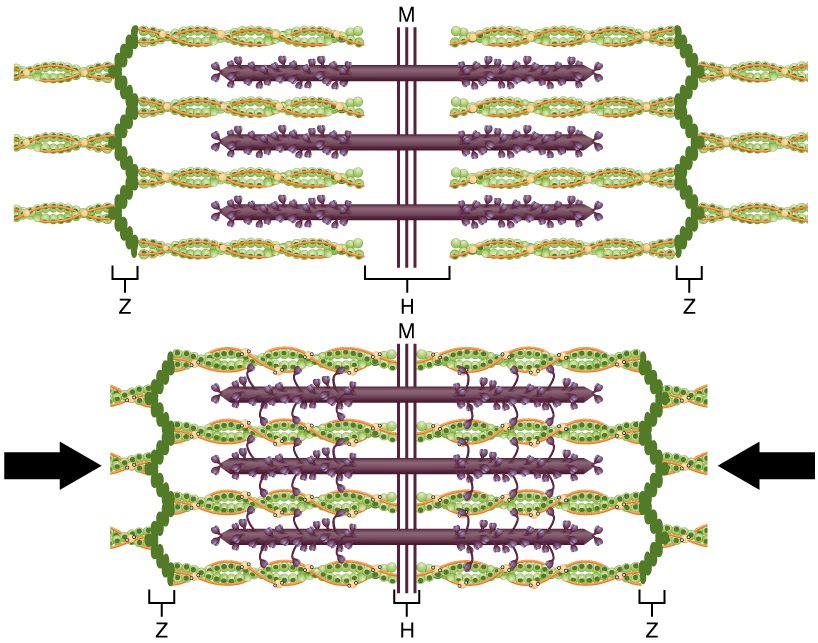
رویدادهای مولکولی مربوط به کوتاه شدن فیبرهای عضلانی در داخل سارکومرها رخ میدهند. انقباض فیبرهای عضلانی مخطط با کوتاه شدن سارکومرها که به صورت خطی در داخل میوفیبریلها آرایش یافته اند، در اثر کشیده شدن سرهای میوزین بر روی اکتینها رخ می دهد.
ناحیهای که فیلامنتهای ضخیم و نازک همپوشانی دارند، دارای ظاهری متراکم است و فضای اندکی بین فیلامنتها وجود دارد. این ناحیه در فرایند انقباض بسیار مهم است و حرکت فیلامنتها در طول یکدیگر از اینجا آغاز میشود. فیلامنتهای نازک یا اکتینها در انتهای خود توسط صفحه Z به یکدیگر متصل شدهاند و به طور کامل به سمت ناحیه مرکزی کشیده نشدهاند. در ناحیه مرکزی سارکومر تنها فیبرهای ضخیم قرار دارند. این فیبرها در خط M از قاعده به یکدیگر متصل شدهاند. هر میوفیبریل از چندین سارکومر تشکیل شده است که در طول آن قرار گرفتهاند. پس میوفیبریلها و سلولهای عضلانی در نتیجه انقباض سارکومرها منقبض میشوند.
مقاله مرتبط: نوروترانسمیتر: تعریف، سیگنالینگ سلولی و انواع
مدل لغزش فیلامنتها در مکانیسم انقباض ماهیچه
پس از دریافت سیگنال نورون حرکتی، فیبر ماهیچه اسکلتی با کشیده شدن فیلامنتهای اکتین به سمت یکدیگر در طول فیلامنتهای میوزین و در داخل سارکومرها، منقبض میشود. این فرایند مکانیسم لغزش فیلامنتها نام دارد وتنها در صورتی رخ میدهد که جایگاههای اتصال میوزین در فیلامنتهای اکتین در معرض اتصال به میوزین قرار بگیرند. تعدادی از رویدادها که آغازگر آنها ورود یونهای کلسیم به داخل سارکوپلاسم است، عامل این پدیده هستند.
تروپومیوزین پروتئینی است که به دور زنجیرههای فیلامنت اکتین میپیچد و از اتصال میوزین و جایگاه اتصال میوزین در اکتین جلوگیری مینماید. اتصال تروپومیوزین به تروپونین تشکیل کمپلکس تروپونین-تروپومیوزین را میدهد که از اتصال سرهای میوزین به نواحی اتصال فعال در فیلامنتهای اکتین جلوگیری میکند. تروپونین همچنین دارای جایگاهی برای اتصال به یونهای کلسیم است.
برای اینکه انقباض عضله آغاز شود، تروپومیوزین باید از روی جایگاه اتصال میوزین برداشته شود و درنتیجه امکان تشکیل پل عرضی میان اکتین و میوزین فراهم گردد. نخستین گام در این فرایند اتصال یونهای کلسیم به مولکولهای تروپونین و سپس کنار رفتن تروپومیوزینها از جایگاه اتصال میوزین است. درنتیجه سرهای میوزین در معرض اتصال به جایگاههای اتصال در اکتین قرار میگیرند و پلهای عرضی تشکیل می شوند. سرهای میوزین سپس فیلامنتهای نازک را در امتداد فیلامنتهای ضخیم و به سمت مرکز سارکومر میکشانند. حرکت ناشی از هر سر بسیار محدود است و پس از رسیدن به محدوده خود باید دوباره به شکل قبلی خود بازگردد. این فرایند نیازمند ATP میباشد.
مقاله مرتبط: گرفتگی ماهیچهای؛ علت و درمان
ATP و مکانیسم انقباض ماهیچه
برای ادامه پیدا کردن لغزش فیلامنتهای نازک در امتداد فیلامنتهای ضخیم، سرهای مولکول میوزین باید به سمت اکتین کشیده شده، به آن متصل شود و به جای قبلی خود بازگردد. این فرایند چندین بار تکرار میگردد و میوزینها به جایگاههای جدیدتری متصل میشوند. این تکرارها، چرخه پل عرضی نامیده شده اند. این حرکت بسیار شبیه حرکت پاروها هنگام پارو زدن یک فرد است: پدال پاروها (مانند سرهای میوزین) به سمت آب کشیده میشود، از آن بیرون میآید و دوباره به موقعیت اصلی خود بازمیگردد. این چرخه چندین بار تکرار میشود. هر چرخه نیازمند انرژی است که توسط ATP تامین میشود.
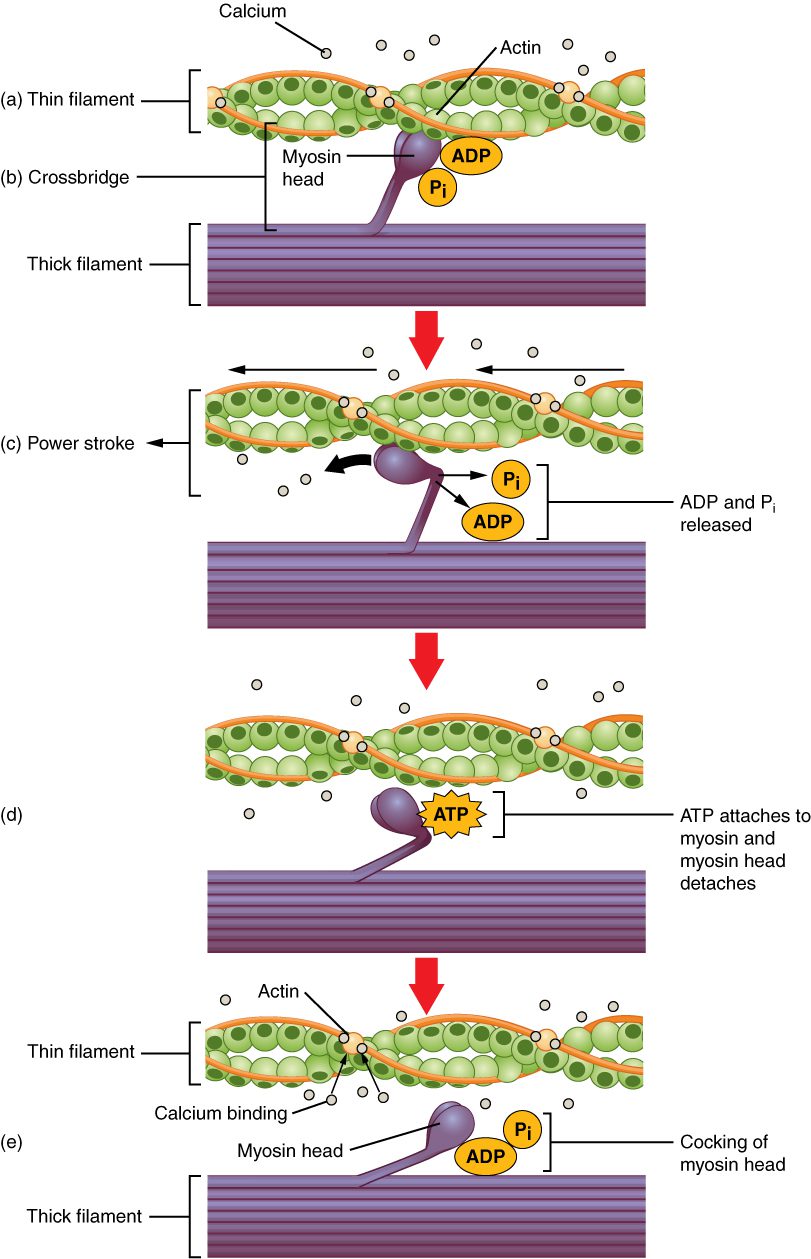
تشکیل پل عرضی زمانی رخ میدهد که سر میوزین به اکتین متصل میشود، در حالیکه ADP و فسفات معدنی همچنان به میوزین متصل هستند. فسفات سپس آزاد شده و باعث اتصال محکمتر میوزین به اکتین میگردد. در مرحله بعدی سر میوزین به سمت خط M کشیده میشود و اکتین را نیز با خود حمل میکند. در این حالت میزان حرکت به سمت خط M در حدود ۱۰ نانومتر است. این حرکت ضربه نیرو نام دارد. در غیاب ATP سر میوزین از اکتین جدا نخواهد شد.
قسمتی از سر میوزین به اکتین متصل میشود و قسمت دیگری از آن جایگاهی برای اتصال به ATP دارد. اتصال ATP باعث جدایی سر میوزین از اکتین میگردد. پس از این اتصال ATP در اثر خاصیت ATPase میوزین به ADP و فسفات آزاد تبدیل میگردد. انرژی آزاد شده در اثر هیدرولیز ATP صرف بازگرداندن سرمیوزین به زاویه خمیده پیشین خود میشود. در این حالت میوزین برای چرخه بعدی آماده است.
هنگامی که سر میوزین به صورت خمیده است، میوزین در بالاترین سطح انرژی خود قرار دارد. این انرژی با حرکت سر میوزین در طی ضربه نیرو آزاد میشود و در پایان آن سر میوزین دارای سطح انرژی پایینی است. پس از ضربه نیرو ADP آزاد می شود. البته در این حالتد پل عرضی همچنان برقرار است و اکتین و میوزین متصل به یکدیگر میباشند. تا زمانی که ATP موجود باشد، به راحتی به میوزین متصل می شود، چرخه پل عرضی ادامه مییابد و عضله منقبض میماند.
لازم به ذکر است که هر فیلامنت ضخیم که حاوی حدود ۳۰۰ مولکول میوزین است، دارای تعداد فراوانی از سرهای میوزین میباشد. در نتیجه حین انقباض عضله پلهای عرضی فراوانی تشکیل شده و از بین میروند. با ضرب این مقدار در تمام سارکومرهای موجود در یک میوفیبریل و تمام میوفیبریلهای موجود در یک فیبر عضلانی و تمام فیبرهای عضلانی یک ماهیچه اسکلتی، میتوان به میزان ATP موردنیاز برای ادامه فعالیت عضلات اسکلتی پی برد. در واقع عدم وجود ATP است که بلافاصله پس از مرگ فرد باعث جمود نعشی میگردد. در صورتیکه امکان تولید ATP بیشتری فراهم نباشد، سر مولکولهای میوزین از جایگاه اتصال اکتین جدا نمیشود و در نتیجه پلهای عرضی برقرار میمانند و سفتی عضلات اسکلتی ایجاد میشود.
سلام آیا کلسیم به میوزین متصل میشود یا نمی شود؟
سلام
خیر؛ کلسیم در عضلهی مخطط به مولکول تروپونین متصل میشود
سلام خسته نباشید منبع این مطالب چیست؟