با افزایش روزافزون توالیهای نوکلئیکاسید شناخته شده و نیز وجود روشهایی برای سنتز الیگونوکلئوتیدهای دلخواه، امروزه میتوان قطعات DNA تکرشتهای کوتاهی را برای انجام هیبریداسیون DNA طراحی کرد. این قطعات پروب DNA نامیده میشوند. پروبها، مولکولهای DNA ای هستند که با موادی همچون ایزوتوپ ها، آنزیمها و کروموفورها، لیبل شدهاند و از طریق جفت شدن با بازهای مکمل و درنتیجه تشکیل پیوند هیدروژنی، میتوانند توالی مکمل خود را شناسایی کنند. پروبهای DNA میتوانند از منابع مختلفی تهیه شوند؛ مانند توالیهای DNA ژنومی تصادفی، برخی ژنها، توالیهای cDNA و یا الیگونوکلئوتیدی که بر اساس دانش ما از توالی آمینواسیدی پروتئینها سنتز شده اند.
مقالات مرتبط:
- هیبریداسیون DNA
- تکنیک FISH
- تکنیک DNA microarray
- تکنیک Southern Blotting
- تکنیک Northern Blotting
به منظور تهیه پروب، توالی و یا ژن شناخته شده ابتدا باید استخراج و به نحوی لیبل شود. شناسایی ژنها امروزه به علت توالییابی شدن بسیاری از ژنومها، به میزان زیادی آسان شده است. به عنوان مثال دانشمندان می توانند نسخهای از یک ژن انسانی را تهیه کرده و به عنوان پروب استفاده کنند تا بتوانند ژنهای مشابه را در سایر ارگانیسم ها شناسایی نمایند. میتوان با استفاده از دادههای حاصل از توالییابی نیز، یک پروب الیگونوکلئوتیدی جدید که بتواند ژن دلخواهمان را تشخیص دهد، طراحی نمود.
لیبل کردن پروبها باعث میشود که بتوانیم بعدا هیبرید تشکیل شده را آشکارسازی کنیم. برای لیبل کردن پروبها فرایندهای متنوعی وجود دارد؛ از جمله، روشهای ایزوتوپی مانند لیبل کردن با ۳۲P و روشهای غیرایزوتوپی مانند استفاده از نوکلئوتیدهای تغییریافته حاوی فلوروفور. تشخیص هیبریداسیون پروب DNAای که به صورت رادیواکتیو لیبل شده باشد، با اتورادیوگرافی ممکن است؛ این در حالی است که قطعات DNA لیبل شده به صورت فلورسنت، در اثر قرار گرفتن در معرض نوری با طول موج معین آشکار میشوند.
با توجه به اینکه توالی نوکلئوتیدی بسیاری از ژنومها شناخته شدهاند، طراحی پروبی که بتواند به ناحیه موردنظر در ژنوم متصل شود، کار دشواری نیست. پروبها تک رشتهای بوده و میتوانند از جنس RNA یا DNA باشند. انواع پرکاربرد پروب معمولا طولی در حدود صدها نوکلئوتید داشته و توالی هدفی را که از لحاظ توالی بسیار به آنها شباهت دارد را تشخیص میدهند. پروبهای الیگونوکلئوتیدی کوتاه طولی در حدود ۳۰ نوکلئوتید دارند (در پروبهای الیگونوکلئوتیدی) و به صورت تجاری نیز سنتز میشوند. کاربرد این پروبها تمایز میان مولکولهای هدفی است که ممکن است تنها در یک نوکلئوتید با هم تفاوت داشته باشند.
احتمال وجود یک توالی ۳۰ نوکلئوتیدی خاص در ژنوم، یک مورد در هر ۱۰۱۸ نوکلئوتید است. با توجه به اینکه ژنوم انسان حاوی ۳ میلیارد جفت نوکلئوتید است، احتمال هیبریداسیون یک پروب ۳۰ نوکلئوتیدی با هر توالی به جز توالی مورد نظر بسیار اندک است. به عبارت دیگر این گونه نیست که توالی مکمل پروب چندین بار در ژنوم تکرار شود. پس شرایط هیبریداسیون میتواند به گونهای تنظیم شود که وجود حتی یک mismatch، از هیبریداسیون توالیهای مشابه جلوگیری کند.
انواع پروبهای DNA
طراحی پروب، بستگی به کاربرد پروب مورداستفاده دارد. پروبها میتوانند ژنی و یا الیگونوکلئوتیدی باشند.
- پروبهای ژنی
پروبهای ژنی طولی بیش از ۵۰۰ باز دارند و کل ژن هدف یا قسمت زیادی از آن را شامل میشوند. این پروبها میتوانند به دو صورت تولید شوند. روش اول جای دادن پروب در ساختار باکتری یا ویروس است؛ به گونهای که با تکثیر باکتری و ویروس، کپیهای فراوانی از پروب تهیه شود. با توجه به اینکه توالی پروب مشخص است میتوان توسط آنزیمهای محدودکننده آن را از بقیه ژنوم جدا کرد.
روش دوم PCR است که ابزاری قدرتمند برای ساخت پروبهای ژنی است؛ چون با استفاده از آن، انجام همزمان تکثیر و لیبلدار کردن قطعات بلند DNA ممکن میگردد. به این صورت که از DNA کروموزومی یا پلاسمیدی به عنوان الگو و از نوکلئوتیدهای لیبلدار برای گسترش قطعه موردنظر استفاده میکنند. اگر کل توالی ژن موردنظر مشخص باشد، پرایمرها میتوانند به منظور تکثیر کل ژن یا قطعاتی از آن مورد استفاده قرار گیرند. در این صورت نیازی به استفاده از آنزیمهای محدودکننده، الکتروفورز و استخراج DNA نخواهد بود. پروبهای ژنی عموما اختصاصیت بالایی نسبت به پروبهای الیگونوکلئوتیدی دارند. علت آن است که توالیشان طولانیتر بوده و قابلیت حمل تعداد بیشتری از گروههای شیمیایی لیبلکننده را دارا میباشند.
- پروبهای الیگونوکلئوتیدی
پروبهای الیگونوکلئوتیدی معمولا برای هدف قرار دادن نقاط خاصی در درون ژن استفاده و به صورت شیمیایی سنتز میشوند. معمولترین این پروبها طولی در حدود ۳۰-۱۸ باز دارند و به صورت تک رشتهای سنتز میشوند. البته تجهیزاتی که امروزه مورد استفاده قرار میگیرند، میتوانند به طور موثری تا طول ۱۰۰ باز را هم سنتز کنند. اگر یک الیگونوکلئوتید توالی شایعی داشته باشد، به بسیاری از ژنهای دیگر نیز متصل خواهد شد. درنتیجه، پروبهای الیگونوکلئوتیدی طولشان باید به گونهای در نظر گرفته شود که دارای جایگاههای اختصاصی اندکی در ژنوم هدف باشند و نیز از هیبرید شدن قطعات شبیه به هم جلوگیری شود. در انتخاب توالی یک پروب الیگونوکلئوتیدی از یک ژن شناختهشده، باید نکات زیر مدنظر قرار گیرند:
- طول پروب باید بین ۱۸ تا ۵۰ باز باشد. پروبهای طولانیتر منجر به افزایش مدت زمان هیبریداسیون شده و انواع کوتاهتر فاقد اختصاصیت هستند.
- در داخل پروب نباید نواحی مکمل هم وجود داشته باشند. وجود این نواحی میتواند منجر به تشکیل ساختارهای موسوم به سنجاق سر (hairpin) شده و هیبرید شدن پروب با توالی موردنظر را مهار کنند.
- پروب نباید حاوی یک رشته طولانی متشکل از تنها یک باز باشد.
- پس از اینکه قطعه موردنظر تولید شد، باید توسط کامپیوتر (in silico) آنالیز شود. توالی پروب باید با توالی ناحیهای که از آن مشتق شده و نیز توالی مکمل آن، مقایسه شود.
نحوه اتصال پروب به توالی مکمل
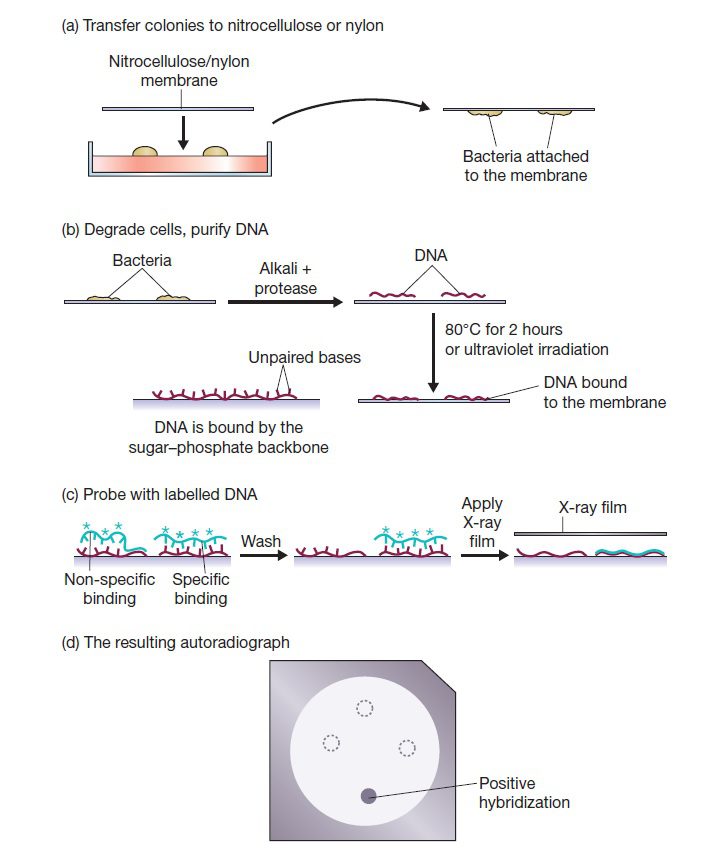
روشی آسان و معمول برای اتصال DNA به پروبهای ژنی و یا الیگونوکلئوتیدی، ثابت کردن نوکلئیک اسید هدف در یک پایه جامد و آزاد بودن پروب در محلول است. جمعیت نوکلئیکاسیدهای موجود در نمونه تست یا به صورت مستقیم خالصسازی و روی پایه رسوب داده میشوند و یا رسوب پس از تفکیک بر اساس اندازه انجام میگیرد. البته سلولها و کروموزومها نیز میتوانند روی پایه ثابت شوند و DNA یا RNA آنها تحت فرایند هیبریداسیون قرار گیرد.
معمولا این پایه میتواند از جنس نیتروسلولز، غشای نایلونی، لاتکس و … باشد. غشاهای نیتروسلولزی معمولا به این منظور استفاده میشوند. البته با اینکه این غشاها توانایی اتصال به تعداد زیادی از قطعات DNA را دارند، اما نمیتوانند قطعات DNA را به صورت محکم در خود نگه دارند و طی هیبریداسیون برخی از قطعات DNA از آنها رها میشود.
پس از جدا کردن قطعات DNA موردنظر توسط الکتروفورز، این قطعات در فیلتر غشایی بیحرکت شده و سپس پروبها به منظور اتصال به توالیهای مکمل خود، افزوده میشوند. پس از تشکیل هترودوپلکسها، پروبهای لیبلدار متصل نشده شسته میشوند. وجود توالی هدف با شناسایی شدن پروب در فیلتر غشایی تایید میشود. از جمله روشهای شناسایی، استفاده از پروبهای لیبل شده با مواد رادیواکتیو است. روشهای مختلفی برای فیکس کردن قطعات روی غشاهای نگهدارنده وجود دارد: Southern blotting، Northern blotting وdot blotting.