در علم نوروساینس، long-term potentiation یا LTP به عنوان پدیده قویکننده فعالیت سیناپس با توجه به الگوی فعالیت اخیر آن میباشد. الگوهای فعالیت سیناپسی شامل مواردی از جمله افزایش دیرپای انتقال سیگنال بین دو نورون میباشد. در طرف مقابل LTP، long-term depression قرار دارد و به عنوان پدیدهای در کاهش فعالیت و قدرت سیناپس شناخته میشود.
بیولوژی سلولیمولکولی حافظه و یادگیری
تحقیقات اخیر با استفاده از مهندسی ژنتیک سعی در شناخت مکانیسمهای درگیر در ایجاد حافظه دارند. این مطالعات در جهت شناخت پیامرسانیهای سلولی و هستهای میباشند که میتواند جهت تسهیل القاء و یا افزایش ثبات شکلپذیری سیناپسی دچار تغییر شوند و لذا اکتساب و نگهداری اطلاعات را تقویت کند. در فرآیند یادگیری و حافظه نقش گیرندههایگلوتاماتی بسیار با اهمیت است.
به طور کلی این گیرندهها به دو دسته یونوتروپیک و متابوتروپیک تقسیم میشوند. ساختار گیرندههای یونوتروپیکی همانند کانال بوده و با فعال شدن، موجب جریان یونهای با بار مثبت به داخل سلول میشود. این دسته از گیرندهها شامل گیرندههای ۱۹AMPA ،۱۸NMDA و کاینات است. فعال شدن گیرندههای متابوتروپیکی گلوتامات ساختار پروتئینی چسبیده به G- پروتئینها را دارند. برخی گیرندههای یونوتروپیک گلوتاماتی، گیرندههای متابوتروپیکی کینتیک فعالسازی و تاثیرگذاری بسیار آهستهای دارند. در نتیجه در فرآیندهای انتقال پایه سیناپسی، تقویت و یا تضعیف سیناپسی گیرندههای یونوتروپیکی نقش مرکزی و اصلی را دارند.
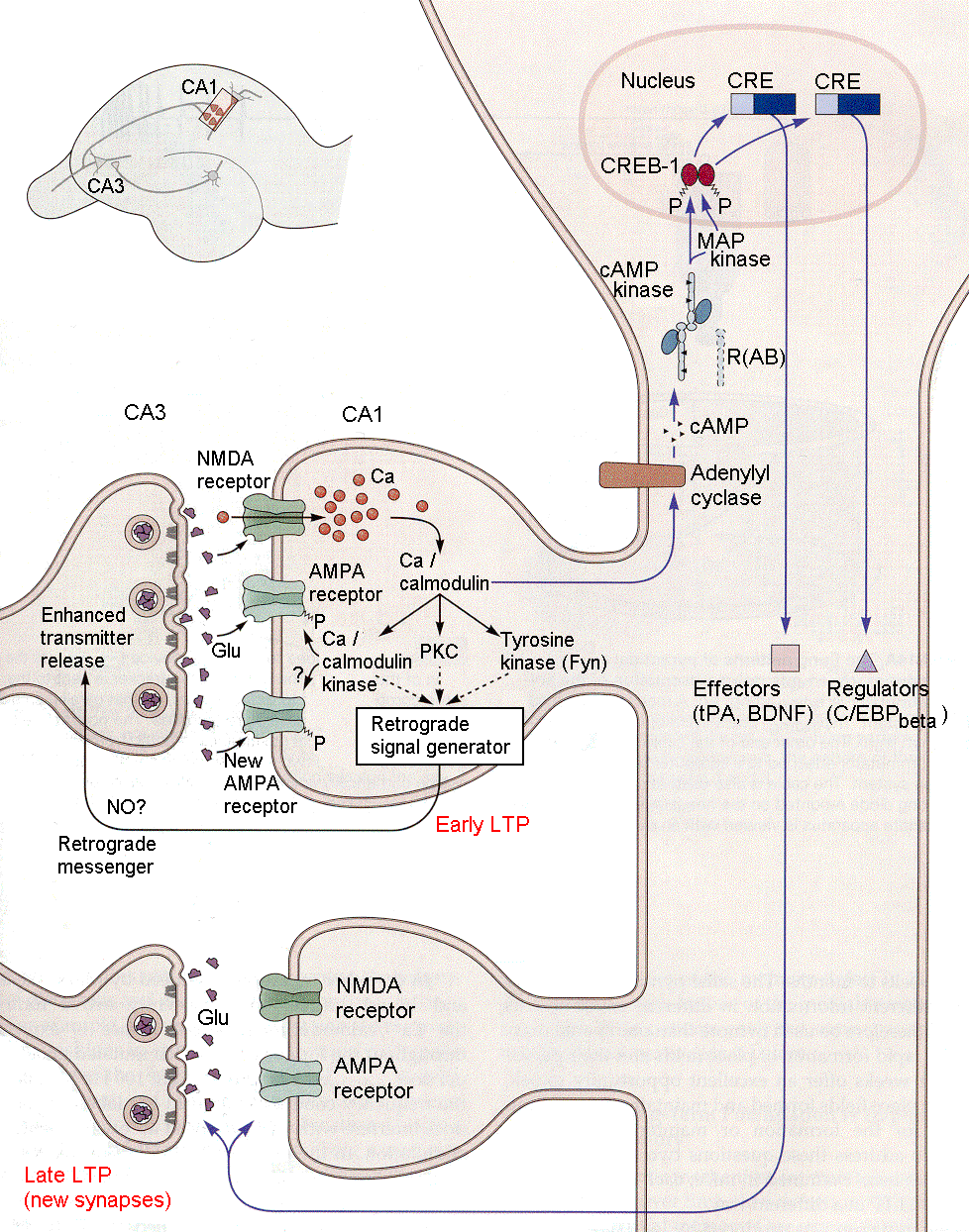 یکی از چندین پدیده کنترل کننده انعطاف سیناپسی (synaptic plasticity) توانایی کنترل قدرت سیناپسهای شیمیایی میباشد. در طول زندگی، مغز ما دائما در حال تغییر است. به این خاصیت تغییر پذیری مغز پلاستیسیته میگویند. به دلیل شباهتش به pastic یا خمیر بازی که میتوان آن را به اشکال متفاوتی درآورد. نه تنها کل مغز، بلکه هر یک از سلولهای عصبی نیز میتوانند به دلایل مختلفی تغییر یابند- مثلا در طول تکامل در سنین پایین، در پاسخ به آسیب مغزی و در حین یادگیری. مکانیسمهای متفاوتی برای پلاستیسیته وجود دارد که مهمترین آنها پلاستیسیته سیناپسی است و عبارت است از علمی که به بررسی قابلیت نورونها برای تغییر تواناییهای خود برای برقراری ارتباط با یکدیگر میپردازد. سیناپس ها به طور کلی به دو نوع الکتریکی و شیمیایی تقسیم میگردند. مطالعات نوروساینتیست ها در سالهای اخیر نشان از ارتباط نزدیک پدیده حافظه و ذخیره اطلاعات در مغز با تغییرات در قدرت سیناپس ها و LTP و LTD دارد. در واقع LTP به عنوان مکانیسم سلولی اصلی در فرآیند یادگیری و حافظه شناخته میشود. مکانیسم LTP در سال ۱۹۶۶ توسط دانشمندی به نام Terje Lomo همزمان با آزمایش بر روی هیپوکامپ موش کشف شد. بخشی از تحقیقات امروزه بر روی این موضوع تمرکز خود را بر مکانیسم بیولوژیکال این پدیده داشته و برخی دیگر به ارتباط این موضوع و یادگیریرفتاری توجه داشته اند.
یکی از چندین پدیده کنترل کننده انعطاف سیناپسی (synaptic plasticity) توانایی کنترل قدرت سیناپسهای شیمیایی میباشد. در طول زندگی، مغز ما دائما در حال تغییر است. به این خاصیت تغییر پذیری مغز پلاستیسیته میگویند. به دلیل شباهتش به pastic یا خمیر بازی که میتوان آن را به اشکال متفاوتی درآورد. نه تنها کل مغز، بلکه هر یک از سلولهای عصبی نیز میتوانند به دلایل مختلفی تغییر یابند- مثلا در طول تکامل در سنین پایین، در پاسخ به آسیب مغزی و در حین یادگیری. مکانیسمهای متفاوتی برای پلاستیسیته وجود دارد که مهمترین آنها پلاستیسیته سیناپسی است و عبارت است از علمی که به بررسی قابلیت نورونها برای تغییر تواناییهای خود برای برقراری ارتباط با یکدیگر میپردازد. سیناپس ها به طور کلی به دو نوع الکتریکی و شیمیایی تقسیم میگردند. مطالعات نوروساینتیست ها در سالهای اخیر نشان از ارتباط نزدیک پدیده حافظه و ذخیره اطلاعات در مغز با تغییرات در قدرت سیناپس ها و LTP و LTD دارد. در واقع LTP به عنوان مکانیسم سلولی اصلی در فرآیند یادگیری و حافظه شناخته میشود. مکانیسم LTP در سال ۱۹۶۶ توسط دانشمندی به نام Terje Lomo همزمان با آزمایش بر روی هیپوکامپ موش کشف شد. بخشی از تحقیقات امروزه بر روی این موضوع تمرکز خود را بر مکانیسم بیولوژیکال این پدیده داشته و برخی دیگر به ارتباط این موضوع و یادگیریرفتاری توجه داشته اند.
مدل و تئوری
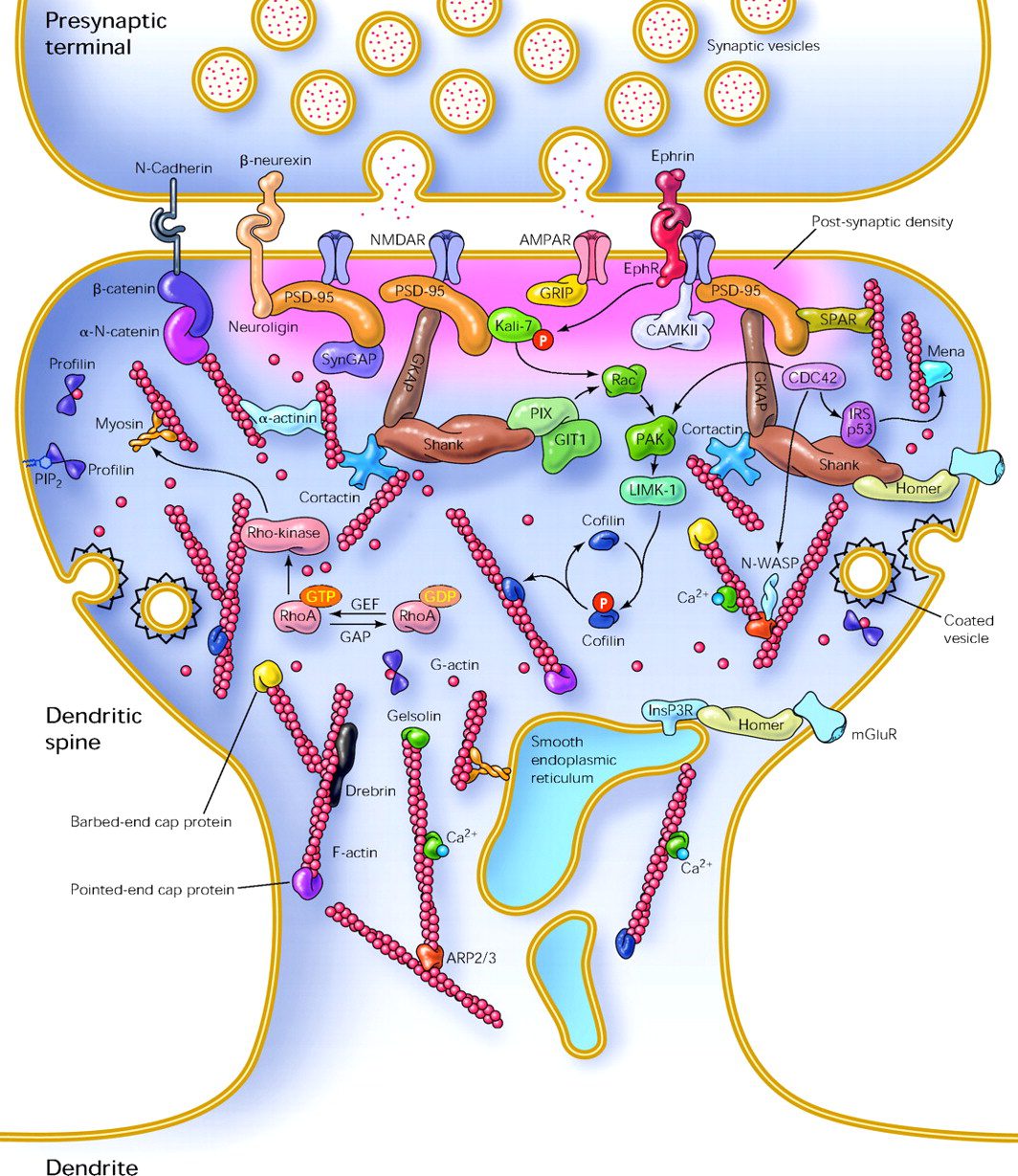 جزئییات فیزیکی و بیولوژیکال این پدیده هنوز به طور کامل درک نشده است. اما مدل های زیادی در این باره مطرح شده است. یکی از مهمترین آن مدل های مرتبط با خارهای دندریتی (dendritic spine) میباشد. این ساختار های متغیر میتوانند در پاسخ به فعالیتهای سیناپسی تغییر ساختار دهند. این تغییرات میتواند در محدوده دقیقه یا ساعت عمل کند. در اینجا ساختار مربوط به خارهای سیناپسی را مشاهده میکنید.
جزئییات فیزیکی و بیولوژیکال این پدیده هنوز به طور کامل درک نشده است. اما مدل های زیادی در این باره مطرح شده است. یکی از مهمترین آن مدل های مرتبط با خارهای دندریتی (dendritic spine) میباشد. این ساختار های متغیر میتوانند در پاسخ به فعالیتهای سیناپسی تغییر ساختار دهند. این تغییرات میتواند در محدوده دقیقه یا ساعت عمل کند. در اینجا ساختار مربوط به خارهای سیناپسی را مشاهده میکنید.
تغییرات صورت گرفته میتواند موجب تغییر در قدرت سیناپسها ایجاد نماید. مدل های ریاضی از جمله تئوری BCM که مرتبط با کلیسم داخل سلولی در ارتباط با گیرندههای ولتاژی NMDA میباشد، از سال ۱۹۸۰ توسعه یافته اند. در حال حاضر عدهای از دانشمندان به ارتباط میان LTP، قدرت سیناپسی و تنظیم گیرنده ها به عنوان مکانیسمهای دخیل در حافظه عقیده دارند.
انواع LTP
از زمان اکتشاف پدیده LTP در هیپوکامپ موش، دانشمندان موفق به کشف کاربرد این پدیده در بخش هایی از جمله قشر مخ، مخچه و آمیگدال مغز شدند. رابرت مالنکا، دانشمند محقق LTP، پیشنهاد میکند که ممکن این پدیده در تمامی سیناپسهای تحریکی پستانداران وجود داشته باشد.
نقاط گوناگون در مغز انواع مختلفی از LTP را نشان میدهند. رخ دادن این پدیده به انواعی از فاکتورها وابسته میباشد. یکی از عوامل سن جاندار میباشد. برای مثال وقوع این پدیده در هیپوکامپ موش نارس عملکری متفاوت نسبت به جاندار بالغ و پیر دارد. این موضوع میتواند نشانه تغییر پتانسیل یادگیری در سنین مختلف باشد.
مسیرهای پیام رسانی مورد استفاده توسط هر سلول نیز در تعیین نوع LTP موثر خواهد بود. برای مثال نوع LTP در هیپوکامپ مرتبط با نوع گیرنده NMDA میباشد. انواع دیگر ممکن است در ارتباط با (metabotropic glutamate receptor (mGluR باشند.
مشخصات
LTP مرتبط با گیرنده های NMDA مشخصات متعددی را نشان میدهند. از جمله ویژه بودن، پیوستگی و همکاری و مقاوت در اینجا به بررسی این موارد میپردازیم.
ویژهبودن
ویژه بودن به معنی عدم گسترش LTP از یک سیناپس به سیناپس دیگر میباشد. LTP تنها به سیناپسهایی با قابلیت مشارکت با سیناپس مورد نظر انتقال مییابد. نکته جالب کاهش فاکتور ویژه بودن در برخی از بیماریها است. نظریهای که به توجیه این مورد میپردازد اولین با در سال ۱۹۹۷ توسط Frey و Morris تحت عنوان synaptic tagging and capture hypothesis انتشار یافت.
پیوستگی
این پدیده در زمان مناسب نبودن محرک ورودی برای تولید LTP مشاهده میشود. تحریکات متناوب قوی در مسیری دیگر مشترک، میتواند سبب ایجاد LTP در هر دو مسیر گردد.
مشارکت
LTP همچنین بوسیله تحریکات تتتانیک مسیری خاص به یک سیناپس نیز ایجاد گردد. اما راه دیگر به کمک تحریکات ضعیفتر از چند مسیر به یک سیناپس میباشد.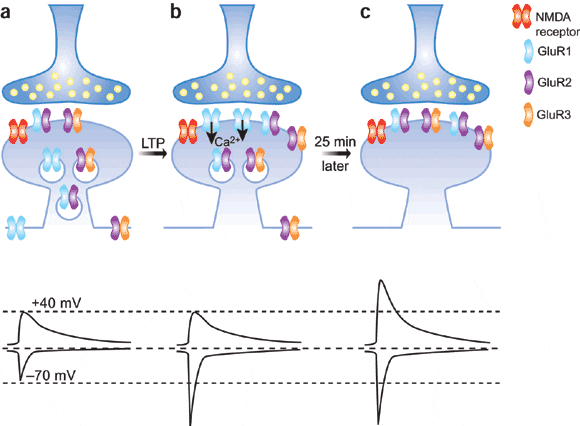
مقاومت
این پدیده میتواند دوامی معادل چند دقیقه تا حتی چندین ماه داشته باشد. این موضوع سبب متمایز ساختن این پدیده از سایر فرآیندهای synaptic plasticity میباشد. تحریک مداوم سیناپس میتواند موجب دوام بیشتر LTP گردد. ارزش این موضوع میتواند در لزوم تکرار و مرور در فرآیند یادگیری مورد بررسی قرار بگیرد.
تعدیل کنندهها و میانجیگرها
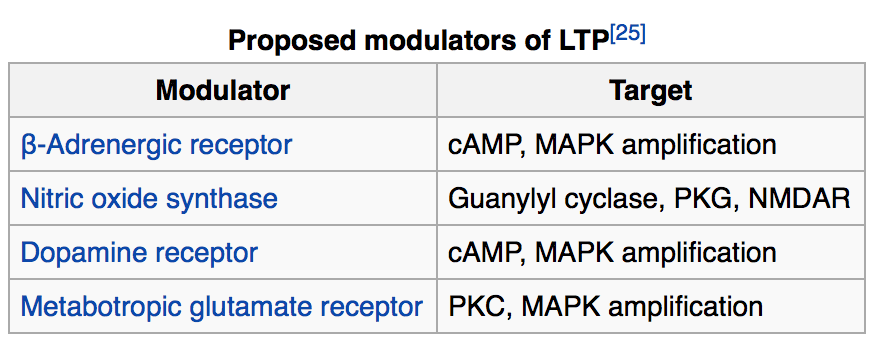 مولکولهایی که موجب ایجاد LTP میگردند در دو دسته میانجیها و تعدیل کنندهها قرار میگیرند. میانجیگر مولکولی مانند گیرنده NMDA ویا کلسیم میباشد که حضور آنها برا ایجاد این پدیده الزامی میباشد. در مقابل تعدیلکننده ها سبب تغییر در کیفیت LTP شده اما برای شروع آن الزامی نمیباشند.
مولکولهایی که موجب ایجاد LTP میگردند در دو دسته میانجیها و تعدیل کنندهها قرار میگیرند. میانجیگر مولکولی مانند گیرنده NMDA ویا کلسیم میباشد که حضور آنها برا ایجاد این پدیده الزامی میباشد. در مقابل تعدیلکننده ها سبب تغییر در کیفیت LTP شده اما برای شروع آن الزامی نمیباشند.
منبع: NCBI | NeuroScience