دیانسفال ناحیهای از لوله عصبی مهره داران است که ساختارهای مغزی خلفی را ایجاد میکند. در روند تکاملی، مغز از پروزنسفالون، قدامیترین وزیکول لوله عصبی ایجاد میشود که خود بعداً به بخشهای دیانسفالون و تلسفالون تقسیم میشود. در بزرگسالان، دیانسفال در انتهای فوقانی ساقه مغز، بین مغز و ساقه مغزی قرار میگیرد و از چهار مؤلفه مجزا تشکیل میشود: تالاموس، ساب تالاموس، هیپوتالاموس و اپی تالاموس.
هیپوتالاموس، ناحیهای از قسمتهای زیرین مغز است که در زیر تالاموس قرار داشته و کف بطن سوم مغزی را تشکیل میدهد. این ساختار مخروطی شکل کوچک، تا ناحیه پایینتر مغز پیش رفته و به وسیله ساقه هیپوفیز به هیپوفیز متصل میشود. هیپوتالاموس مرکز کنترل بسیاری از فعالیتهای سیستم عصبی اتونوم است و به دلیل تعاملی که با غده هیپوفیز دارد، سیستم غدد درون ریز بدن را نیز کنترل میکند. از جمله وظایف هیپوتالاموس میتوان به موارد زیر اشاره کرد:
- ترشح هورمون
- تنظیم دمای بدن
- کنترل چرخههای فیزیولوژیکی روزانه
- کنترل اشتها
- مدیریت رفتارهای جنسی
- تنظیم پاسخهای عاطفی
پژوهشها و کارهایی که والتر رودولف هس (walter rudolf hess) انجام داده است، بخش بزرگی از دانش کنونی ما از هیپوتالاموس و عملکردش را تشکیل میدهد و چراغ راه محققیان پس از شد. جایزه نوبل سال ۱۹۴۹ مشترکاً به والتر رادولف هس به پاس تحقیقاتش در رابطه با دیانسفالون و آنتونیو اگاس مونیز (Antônio Egas Moniz) به پاس کشف اهمیت درمانی لوبتومی فرونتال تعلق گرفت. هیس با انجام آزمایش بر روی گربهها و سگها، دریافت که دیانسفال، بخصوص بخش هیپوتالاموس، در کنترل و هماهنگی عملکرد خود مختار نقش دارد. در واقع او مناطق خاص و کوچکی از هیپوتالاموس را تحریک الکتریکی کرده و مشاهده کرد که با این کار، بسته به منطقه تحریک شده، ضربان قلب، فشار خون و تنفس حیوانات در حالتی مانند ترس (واکنش سمپاتیک) افزایش مییابد. علاوه بر این تحریک نقطه دیگر باعث کاهش ضربان قلب، فشار خون، تنفس و افزایش فعالیت دستگاه گوارش (واکنش پاراسمپاتیک) میشود.
والتر رودولف هس
والتر ردولف هس در سال ۱۸۸۱ در فرآنفلد (Frauenfeld) سوئیس متولد شد. با تشویقهای پدرش به علم علاقه مند شد و به همراه او به انجام آزمایش در آزمایشگاه فیزیک پرداخت. تحصیلات پزشکی خود را در سال ۱۸۹۹ در لوزان (Lausanne) آغاز و سپس در برلین (Berlin)، کیل (Kiel) و زوریخ (Zurich) ادامه داد. در سال ۱۹۰۶ موفق به اخذ مدرک پزشکی از دانشگاه زوریخ شد و تحت نظر کنراد برونر (Conrad Brunner) به یادگیری جراحی پرداخت. وی موفق به اختراع دستگاه ویسکوزیمتر به منظور اندازه گیری ویسکوزیته خون شد و نتیجه مطالعات خود را در سال ۱۹۰۶ منتشر کرد. در سال ۱۹۰۷، به دانشگاه زوریخ رفت تا زیر نظر اتو هاب (Otto Haab) به عنوان یک چشم پزشک آموزش ببیند و مطب شخصی خود را در Rapperswil SG باز کند.
هس در این سالها علاوه بر تشخیص بیماریهای چشمی، مهارتهای جراحی بسیاری را آموخت که بعدها در انجام آزمایشاتهای انجام بر روی حیوانات به او کمک زیادی کرد. در این سالها، او موفق به کشف صفحه نمایش هس یا دستگاه هماهنگ سنج شد. او به طور مستقل سیستمی را طراحی کرد که با استفاده از آن بتواند عضلات چشمی فلج شده در دیپلوپیا و انحراف چشمی را تشخیص دهد. از این دستگاه هنوز هم در حوزه چشم پزشکی امروزی استفاده میشود.
او در سال ۱۹۱۲، با وجود هشدارهای همسرش لوئیز در مورد مشکلات مالی، شغل خود را رها کرد و تحت نظر جاستوس گول (Justus Gaule) به پژوهش پرداخت و در سال ۱۹۱۳ به عنوان دانشیار غیر رسمی آغاز به کار کرد. هدف اصلی تحقیقات وی، بررسی تنظیم جریان خون وسیستم تنفسی بود. در طول جنگ جهانی اول نیز یک سال را در انستیتوی فیزیولوژیک دانشگاه بن تحت نظر مکس ورورن (Max Verworn) گذراند. در سال ۱۹۱۶، هس به عنوان مدیر موقت گروه انستیتوی فیزیولوژیک در دانشگاه زوریخ انتخاب شد و در سال ۱۹۱۷ به طور رسمی به جایگاه استادی و مدیر کلی این موسسه ارتقاء یافت تا زمان بازنشستگی خود در سال ۱۹۵۱ در این جایگاه مشغول خدمت بود.

به عنوان مدیر موسسه، هس میتوانست حیطه مورد نظر خود را برای تحقیق انتخاب کند و در همین راستا از حدود سال ۱۹۲۹ او به تحقیق بر روی پدیدههای داخلی و به خصوص بیولوژی خواب علاقه مند شد. او پژوهش بر روی ناحیه داینسفالون یا بین مغزی را با تهیه نقشهای از قسمتهای مختلف دیانسفالون در گربهها، که در کنترل فرآیندهای داخلی بدن نقش دارند، آغاز کرد. به واسطه همین تحقیقات او در سال ۱۹۴۹ موفق به کسب جایزه نوبل پزشکی و فیزیولوژی شد. با این که کاربرد تحقیقات پایهای وی در روشهای درمانی برای هس مهم نبود اما با این وجود، روش تحریک الکتریکی عمقی مغز، مدتهاست که در درمان علائم بیماری پارکینسون و سایر اختلالات حرکتی کاربردهای زیادی داشته است.
هس در سال ۱۹۵۱ بازنشسته شد اما به کار در دفتر خود در دانشگاه ادامه داد. در سال ۱۹۶۷ ، به آسكونا (Ascona) رفت و در سال ۱۹۷۳ در سن ۹۲ سالگی به علت نارسایی قلبی در لوكارنو (Locarno) سوئیس درگذشت.
پژوهشهای هس درباره هیپوتالاموس
والتر رادولف هس با مطالعه کارهای Karplus & Kreidl در سال ۱۹۱۰و پژوهشهای بارد (Bard) در مورد ارتباط سیستم عصبی سمپاتیک و هیپوتالاموس، به مطالعه دیانسفال علاقه مند شد. وجود گیرندههای مختلف از جمله گیرندههای گرما و اسمورسپتورها، همگرایی ورودیهای احشایی، چشایی و بویایی، وجود خروجیهایی به هیپوفیز و سیستم قدرتمند خروجی حرکتی خارج هرمی* و اتونوم نشان میدهد که ناحیه دیانسفال نقش مهمی را در فرآیندهای تنظیمی بدن ایفا میکند. البته لازم به ذکر است که هیس تنها کسی نبود که به بررسی دیانسفال و نقش آن در کنترل مکانیسمهای داخلی پرداخته بود، بلکه نتیجه گیریهای مشابهی نیز توسط هم عصران هس از جمله رانسون (S.W. Ranson) مطرح شده بود.
تکنیک تحریکی مورد استفاده هس
در اواخر دهه ۱۹۲۰ از روشهای قدیمی تحریک الکتریکی مغز که توسط Fritsch & Hitzig در سال ۱۸۷۰ ابداع شده بود، استفاده میشد. اما هدف اصلی هس استفاده از محرکی بود که تا حد ممکن به محرکهای طبیعی شبیه باشد و بتوان تحریک مغز را در حالت هوشیاری حیوانی که در حال حرکت است، انجام داد. با این روش او میتوانست شبکههای عصبی در حال رشد* تالاموس، هیپوتالاموس و مناطق مجاور مغز میانی و تلنسفالون را بررسی کند.
هدف هس تحریک الیاف كوچک سیستم خودمختار بود. این الیاف، به خصوص در ناحیه خاکستری اطراف بطنها، یا میلینه نشده یا اندک میلیه شده هستند. همچنین او در صدد بود تا این کار با حداقل تحریک یا عدم تحریک الیاف ضخیم میلینه انجام شود تا اثرات محدود کننده بر روی سیستم اتونوم به حداقل برسد.
اولین مرحله ابداع یک قطعه جدید برای تحریک عمقی مغز گربه بود. رادولف هس به منظور به حداقل رساندن* و اختصاصی شدن ناحیه مورد تحریک، کوچکترین الکترود ممکن از جنس استیل ضد زنگ را تهیه کرد. سپس، یک سری آزمایشهای سیستماتیک انجام شد تا تحریکاتی با فرکانس پایین، پالسهای جریانی مستقیم و به شکل موجهای ویژهای انجام شوند. این جریانها حداقل آستانه و پراکندگی جریان را دارند، بنابراین مناسبترین نوع موج، برای تحریک ناحیه خاکستری اطراف بطنها محسوب میشدند.*
هس از تکنیک ویژه تحریک جریان مستقیم (DC) استفاده کرد. او به جای تحریکات موج کوتاه فارادی، از محرکهای طولانی اثرتر ۲۵ میلی ثانیهای* استفاده کرد و برای جلوگیری از اثرات قطبی شدن که ممکن بود باعث آسیب بافتهای مجاور الکترودها شود، از یک “ضد جریان” بسیار ضعیف بین محرکها استفاده کرد تا بار الکتریکی خالص جمع شده را خنثی کند.
موجهای تحریکی به طور معمول ۳۰ ثانیه یا یک دقیقه طول میکشیدند و هر دو نوع تحریک دو قطبی و تک قطبی قابل استفاده بود. علاوه بر این، شدت محرکها نیز ضعیف بود (در حدود ۰.۵-۱.۵ ولت) و فرکانس پایینی در حد ۲–۱۲ هرتز (معمولاً ۸ هرتز) داشتند. برای محدود نگه داشتن گسترش جریانهای تحریکی، که احتمالاً ۰.۱ تا ۰.۱۵ میلی آمپر بودند، دقت زیادی شد. هس از الکترودهای بسیار ظریفتری با قطر ۰.۲۵ میلی متر استفاده کرد. پس از اتخاذ تمامی روشها و کنترلهای بالا هس تلاشهای کرد تا محل آناتومیک دقیق تحریکها را مشخص کند.
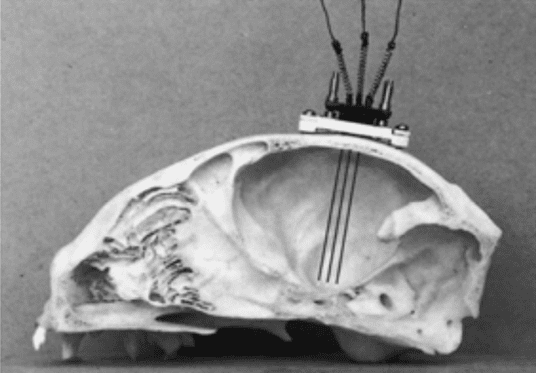
هس از الکتروکواگولاسیون به وسیله الکترودهای تحریکی به منظور محلی سازی و در عین حال تولید ضایعات كوچك استفاده کرد. بدین طریق، امكان مشاهده تغییرات رفتاری و الیاف عصبی تخریب شده ناشی از ضایعات فراهم شد.
رویکرد جدید، منجر به ایجاد نتایجی دقیق و شناخت شدن هس و روش مورد استفادهاش شد. جای گذاری درست و دقیق الکترودها و محلی سازی نقاط تحریک شده، موجب شد تا هس تدوین اطلس بافت شناسی خود بر اساس برشهای بافت شناسی در سه صفحه اصلی داینسفال گربه کرد را آغاز کند.
سیستم اوتونوم
حیوانات هیجانات خود را بسته به شدت محرک، با رفتارهای متفاوتی نشان میدهند. قابل توجهترین واکنش حیوانات، واکنش تحریک با حداقل آستانه تحریک است* که بر روی ناحیه پریفورنیکال اعمال میشود؛ مانند حالتی که گربه یک سگ را میبیند. هیجان شدید حیوانات با خزیدن، تکان دادن دم، جمع شدن گوشها، گشاد شدن مردمک، سیخ شدن موهای پشت کمر و دم، صدا کردن* و غریدن نشان داده میشود. به ندرت ممکن است واکنش ادرار کردن نیز مشاهده شود. در نهایت با تحریک بیشتر، رفتارهای دفاعی مانند پرواز کردن* یا حمله مستقیم بروز مییابد؛ به عنوان مثال گربه به طرف ناظران حمله میکند.

نکتهای که در این مطالعات حائز اهمیت است، اوج تحریک “واکنش دفاعی عاطفی” است و با قطع شدن محرک نیز این واکنش به صورت ناگهانی خاموش میشود. پاسخهای رفتاری پیچیده دیگر، که منشا آنها از مناطق مختلف هیپوتالاموس و سپتوم* است، در هر گونه به شیوه خاص خود الگوهای گرسنگی و تشنگی، حرکات * ، دفع ادرار و مدفوع بروز مییابد.
برای هس این سوال مطرح شد که آیا این واکنشهای آزمایشی صرفاً ناشی از تحریک الکتریکی ناخودآگاه هستند یا این كه به رفتارهای انگیزشی آگاهانه مرتبط هستند. به عقیده وی، پاسخهای رفتاری کاملاً تلفیقی از هماهنگی کامل بین تغییرات حرکتی و خود مختار با درک حیوان از محیط بودند. *
هنس منطقه مربوط به هیپوتالاموس خلفی شکمی را “منطقه ارگوتروپیک ” نامید. با تحریک ناحیه اطراف منطقه کانونی پریفورنیکال علائم واضح هیجان عمومی مانند ناآرامی حرکتی، گشادی مردمک، تنفس سریع و گاهاً حرکات دفاعی بی نتیجه مشاهده شد. هس هنگام تحریک این نواحی در حالت بیهوشی حیوان، شاهد افزایش سریع فشار خون و تنفس بود. محققان بعدی برانگیختگی قشر مغزی را نیز در EEG مشاهده کردند.
منطقه “تروفوتروپیک” در ناحیه قدامیتر در هیپوتالاموس قرار داشته و تا منطقه سپتوم را نیز در بر گرفته است. اثرات تحریک الکتریکی در آن منطقه به طور معمول به سیستم عصبی پارا سمپاتیک نسبت داده میشود. با تجریک این نواحی تظاهرات زیر مشاهده شد: افت فشار خونتوسط که احتمالاً به فعالیت سینوس کاروتید نسبت داده شده، کند شدن تنفس، تنگی مردمک و گاهاً مهار سیستم قلبی. (عکس ۸). در حیوانات هوشیار تغییرات مشابه به صورت آرام شدن رفتار حیوان، شل شدن عضلات و در نهایت به بروز حالت خواب الودگی مشهود بود. محققان دیگر نشان دادند که ضایعات موجود در این منطقه، باعث ناتوانیهای حیاتی، مانند ادم ریوی و هایپر ترمی یا افزایش دمای بدن میشود که مرگ بیمار در این حالت بسیار محتمل است.
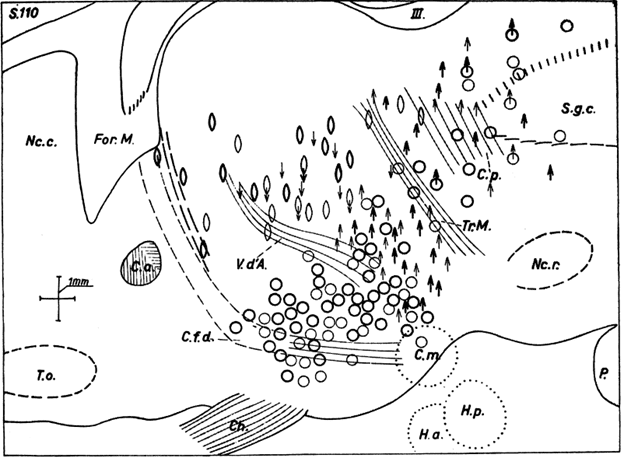
ناحیه تروفوتروپیک تا حدودی با با ناحیه مربوط به خواب تالاموس هم پوشانی داشته داده شده و با تنگی مردمک (بیضیها) و کاهش حرکت (فلشهای رو به پایین) همراه است.
مطالعات مربوط به خواب
هس با مشاهده تغییرات مختلف مربوط به حالت خواب نتیجه گرفت که مناطق خاصی از مغز در هنگام خواب فعال هستند. هس با استفاده از تحریک الکتریکی دیانسفال و مناطق اطراف، متوجه شد که مناطقی که باعث القای خواب میشوند و مناطقی که باعث رفع حالت خواب آلودگی میشوند، هر دو، قادر به تغییر در الگوی خواب و برانگیختگی(aurosal) هستند.
یافتههای هس در رابطه با فرآیند خواب از همان ابتدا بحث برانگیز بود و سالها نیز نقطه بحث پژوهشگران بود. زیرا آنها نتوانستند با تحریک تالاموس داخلی باعث القای خواب در حیوان شوند. پسر والتر رودولف هس، رودولف مکس هس، که الکتروانسفالوگرافی را به سوئیس معرفی کرد، با همکاری کنارد آکرت (Konrad Akert) و ورنر کولا (Werner Koella ) آزمایشهای پدرش را با استفاده از همان محرکها در سال ۱۹۵۰ تکرار کرد و توانست الگوهای معمول خواب EEG را ثبت نماید. شاید علت عدم موفقیت محققان مخالف، این بود که از محرکهای معمولی فارادی یا جریانهای متناوب با فرکانس میانه استفاده میکردند و این محرکها برای تحریک فیبرهای عصبی بدون میلین مناسب نبودند.
علاوه بر این، به دلیل تاخیری که بین آغاز تحریک و خوابیدن حیوان وجود داشت، آنها این فرضیه مطرح میکردند که گربههای هس تنها به علت احساس راحتی که داشتند میخوابیدند. با این حال، هس معتقد بود که گربهها پیش از خوابیدن خود را جمع میکنند و این تأیید میکند که خواب القایی توسط او فیزیولوژیک است. علاوه بر این گربهها میتوانند بلافاصله از خواب بیدار شوند. هس اظهار داشت که پاسخ آهسته به محرک ویژگی تحریک تروفوتروپیک است. وی توضیح میدهد که گربهها در طول زمان تحریک، که حدود ۳۰ تا ۶۰ ثانیه به طول میانجامد، نمیخوابند زیرا احتمالاً پیامی مبنی بر بیدار بودن از طریق تحریک همزمان سایر سیستمها انتقال داده میشود زیرا گاهی همراه با تحریک، میوکلونوس خفیفی نیز به طور همزمان مشاهده میشد.
نکته جالب این جاست که هنگامی که هس شدت محرک را به بیش از ۱.۵ V افزایش میداد، اثر متضادی را (برانگیختگی) مشاهده میکرد. این ممکن است بدین معنی باشد که شبکههای مختلفی با اثرات متناقض و متفاوت بر روی هوشیاری و خواب در بخش تالاموس داخلی (هستههای میانی / سیستم اینترالامینار * بخش خاکستری مرکزی) وجود دارند و شاید این بتوانند توجیه کننده علت نتایج ناسازگار میان محققان باشد. مطابق این فرضیه، ضایعات موجود در تالاموس داخلی، بسته به محل دقیق ضایعه، نورونهای مختلف با تعداد متغیر را تحت تأثیر قرار میدهند. در نتیجه ممکن است حالتهای زیر یا ترکیبی از هر دو مشاهده شوند: کاهش برانگیختگی (کاهش هوشیاری، هایپرسومنیا یا خواب بیش از حد) یا کمبود خواب (بی خوابی).
سیستم موتور
در مورد مکانیسم عملکرد حرکتی، هس در دیانسفالون هماهنگی فضایی-زمانی حرکات سر و بدن در سه محور اصلی را نشان داد. او به تاثیر متقابل سیستمهای حرکتی خلفی (اکستراپیرامیدال) و سیستم هدف مدار تاکید داشت و معتقد بود که تاثیر حرکتی دیانسفال جزئی از سیستم یکپارچه حرکتی است. او اظهار داشت که پوسچر یا وضعیت (Posture) نه تنها حرکات هدف محور را تثبیت و پشتیبانی میکند، بلکه شرایط اولیه لازم را برای انجام فعالیتهای اختیاری را با ارائه یک مدل کنترلی پیش بینی کننده، فراهم میکند.
پژوهشهای هس موجب پیشرفتهای چشمگیری در حوزه علوم عصبی شد و توانست شکاف بین فیزیولوژی و علوم شناختی را به نوبه خود پر کند و پایههایی از دانش کنونی ما در مورد سیستم عصبی مرکزی را ایجاد کرد. همچنین اطلسی که او به روش محلی سازی عملکردی از دیانسفال تهیه کرد، منجر به انجام جراحیهای موفقیت آمیز برای درمان برخی از اختلالات حرکتی در انسان شد.