ساختار DNA که در سال ۱۹۵۳ توسط واتسون و کریک کشف شد، یک دستاورد تاریخی بود. قسمت کلیدی کشف آنها، ساختار مارپیچ دورشتهای DNA و کنار هم قرارگرفتن این دو رشته در اثر پیوندهایی بین جفتبازها بود. این جفتبازها امروزه با عنوان Watson-crick pairs شناخته میشوند: آدنین با تیمین و سیتوزین با گوانین جفت میشود. بازهای DNA به صورت توتومر (ایزومرهایی که به آسانی به همدیگر تبدیل میشوند، این تبدیل غالبا با جابهجایی پروتون صورت میگیرد) هستند و در جفتبازهای A.T و G.C، هر باز به فرم توتومری غالب خود موجود است. نحوه گرد آمدن اجزای DNA کنار هم در آن زمان مشخص نبود، اما کشف جفتبازها مشخص کرد که توالی نوکلئوتیدهای یک رشته، توالی رشته مکمل را مشخص میکند.
کشف ساختار DNA به دستیابی به نتایج دیگری نیز منجر شد. به عنوان مثال مدلی را برای توضیح جهشهایی که ناخواسته هنگام همانندسازی در DNA ایجاد میشوند، ارائه داد. واتسون و کریک احتمال دادند که علت رخ دادن جهشها، حضور یکی از بازها به شکل توتومری غیرمغلوب خود، دقیقا همزمان با ساخت رشته مکمل است. به عبارت دیگر، در صورت حضور بازها در فرم توتومری نامطلوب خود، جفت شدن نادرست بازها و تولید A.C و G.T رخ میدهد. این جهشها از آنجا که ساختار DNA را به هم نمیزنند، به آسانی در جای خود تثبیت میشوند. این نوع از جهشزایی، علیرغم نبود شواهد تجربی توسط بیولوژیستها پذیرفته و در کتابها وارد شد.
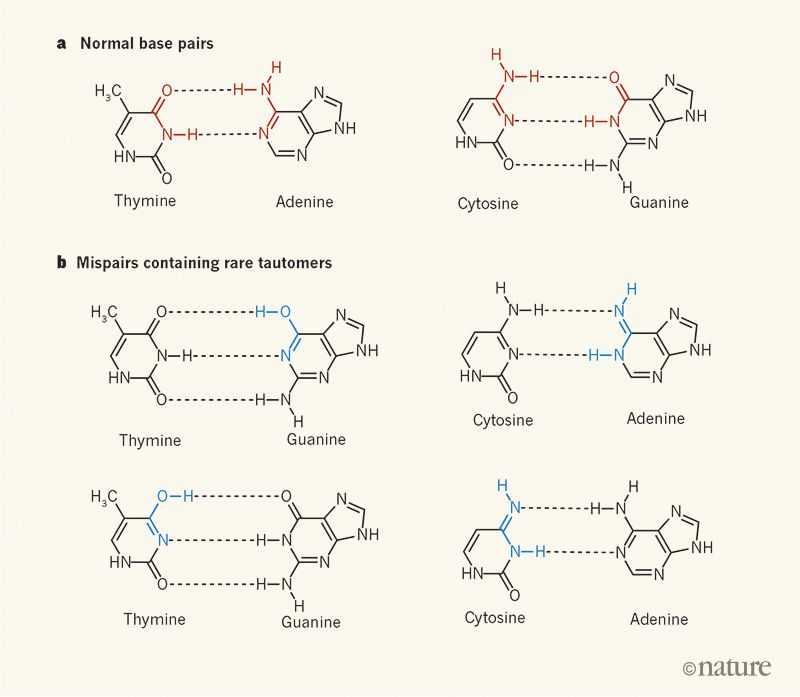
سایر ساختارهای درست جفت نشده (mispairs) علاوه بر ساختارهای حاصل از توتومریزاسیون، در اواسط دهه ۶۰ میلادی کشف شدند. این ساختارها شامل جفتبازهای wobble و Hoogsteen بودند. در اواسط دهه ۸۰، mispairهای مرتبط با انواع باردار DNA نیز شناسایی شدند. در سال ۲۰۱۱، با بررسی کریستالهای DNA با اشعه X، جفتباز C.A که به علت حضور یک توتومر نادر رخ دادهبود، مشاهده شد. این جفت شدن نادرست بازها ، بین نوکلئوتیدهایی که به جایگاههای فعال DNA پلیمراز (آنزیمی که در سنتز DNA از نوکلئوتیدها نقش دارد) متصل بوده و در حضور یون منیزیم، شکل گرفتهبودند. گفته میشود که این یونها جهشزا هستند. ساختار دومی که در همان سال گزارش شد، جفتباز G.T یونیزه شده بود که بین سوبستراهای متصل به DNA پلیمراز شکل گرفته بود. هر دوی این موارد، ساختار مشابهی با mispairهای گزارش شده توسط واتسون و کریک داشتند.
مقاله مرتبط: مولکول DNA پیچخورده میتواند محل جهش خود را نمایان کند
در مدل جهشزایی واتسون و کریک، جفت شدنهای نادرست مرتبط با فرمهای توتومری نامطلوب، بستگی به تناوب دفعاتی دارد که DNA پلیمراز جفت کردن را نادرست انجام میدهد. این فرکانس از یک در هزار تا یک در میلیون متغیر است. تصور میشد که این شکلهای توتومری و جفتبازهای مرتبط با آنها در شکلهای دو رشتهای DNA مشاهده نمیشوند. اما در سال ۲۰۱۵، ۶۵ سال پس از ارائه شدن مدل واتسون و کریک برای جهشزایی، محققان دستاوردی بزرگ را گزارش کردند: آنان با استفاده از طیفسنجی NMR (nuclear magnetic resonounce)، G.Tهای wobble با عمر طولانی را شناسایی کردند. در این ساختارهای mispair ناپایدار و مرتبط با فرمهای توتومری G.T، با جفت شدنهای نادرست حاصل ازG.T های یونیزه در تعادلی پویا قرار داشتند.
نخستین گام در تشکیل جفتباز G.T، اتصال dGTP (نوکلئوتید حاوی باز G) به جایگاه فعال DNA پلیمراز است. سپس آنزیم فعالیت کاتالیزوری خود را انجام داده و DNA پلیمراز پس از تشکیل G.T به فعالیت خود ادامه میدهد. هنگامی که dGTP به جایگاه فعال متصل میشود، پیوند تشکیل شده بین dGTP و T از رشته مقابل، یک ساختار wobble را ایجاد میکند. اما به نظر میرسید که نمیتواند تغییر شکل مورد نیاز برای انجام شدن فعالیت آنزیمی را به دست آورد.
مطالعه اخیر Kimsey و همکارانش، با جمعبندی اطلاعات به دست آمده از آنالیز ساختاری جفتبازهای G.T در ساختار دو رشتهای DNA با سنجش فعالیتهای آنزیمی DNA پلیمراز و مدلهای کامپیوتری، نشان دادند که توتومریسم قطعا عامل جفت شدن نادرست بازها ست. DNA پلیمراز برای اینکه مطمئن شود وظیفهاش به درستی انجام شده، نیاز به تطبیق هندسی نوکلئوتیدها با جایگاه فعال دارد. این تطبیق در مورد G.Tها زمانی رخ میدهد که به صورت یکی از ساختارهای واتسون-کریک (شکل توتومری غیرمغلوب یا ساختار یونیزهشده) باشند. Kimsey و همکارانش به این نتیجه رسیدند که در pH خنثی، حداقل ۹۹ درصد mispairهای G.T مربوط به فرمهای توتومری و نه فرم یونیزه است.
مقاله مرتبط: کشف جهش ژنی جدید مرتبط با ترمیم DNA و آنمی فانکونی
با تعیین موفقیتآمیز نقش فرمهای توتومری در تشکیل G.Tها توسط Kimsey و همکارانش پاسخ نیمی از مسئله جهشزایی ناخواسته به دست آمده است. نیمه دیگر، نیازمند شناسایی فرمهای توتومری C.A مرتبط با جفت شدن نادرست بازها و در ساختار DNA دو رشتهای میباشد. تاکنون NMR و اشعه X موفق به شناسایی ساختار C.A یونیزه در DNA دورشتهای شدهاند.
چالش جدید دانشمندان، کشف مکانیسم جهشزایی ۲-آمینوپورین است. این باز آنالوگ آدنین و گوانین بوده و جهشزا میباشد. به عنوان مثال، این باز جهشزای بالقوهای در باکتریوفاژ T4 است و تعداد جهشهای تبدیل کننده A.T به G.C و برعکس را به میزان ۵۰ برابر بیشتر از جهشهای توضیح داده شده افزایش میدهد.