نوروترانسمیتر های آمینواسیدی بخش اعظم نرورترانسمیترهای مهاری و تحریکی سیستم عصبی را تشکیل می دهند. ارتباط نورون حسی به حرکتی در نخاع که رفلکس پرش زانو را کنترل می کند٬ نقطه آغازی بسیار خوب برای توضیح این مطلب است.
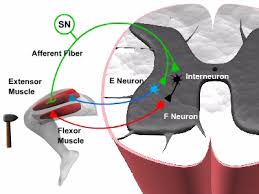
در تصویر یک ارتباط مونوسیناپسی را در نخاع مشاهده می کنید. این ارتباط بین نورون حسی (سبز رنگ) و نورون حرکتی که عضلات اکستانسور را عصبدهی می کند (آبی رنگ)٬ می باشد. پتانسیل عملی که نورون حسی را برانگیخته است باعث ایجاد پتانسیل پس سیانپسی تحریکی (EPSP) در حدود ۱mv در نورون حرکتی اکستانسور می شود. همان نورون حسی با یک نورون بینابینی (سیاه رنگ) نیز سیناپس برقرار می کند. این نورون بینابینی نیز با یک نورون حرکتی دیگر ( قرمز رنگ) که عضله فلکسور را عصبدهی می کند٬سیناپس می دهد. پتانسیل عمل ایجاد شده در نورون بینابینی ٬ باعث ایجاد پتانسیل مهاری پس سیناپسی (IPSP) در نورون حرکتی فلکسور می شود.
به یاد داشته باشید که : پتانسیل های تحریکی پس سیناپسی (EPSP) زیادی مورد نیاز هستند تا پتانسیل استراحت نورون حرکتی به حدی برسد که پتانسیل عمل ایجاد شود.
نوروترانسمیتر هایی که در فرایند های ذکر شده و پاسخ های مهاری یا تحریکی دیگر میانجیگری می کنند ٬ موضوع اصلی این مطلب هستند .
انتقال پیام های تحریکی (تولید EPSP ها) به طور عمده توسط آمینواسید گلوتامات و انتقال عصبی مهاری (IPSP ها) عمدتا توسط گلیسین در نخاع و در مغز توسط یک متابولیت گلوتامات به نام گاما آمینوبوتیریک اسید یا گابا (GABA) انجام می گیرد.
تفاوت فضایی و ریخت شناسی (مورفولوژیکال) بین ورودی های تحریکی و مهاری
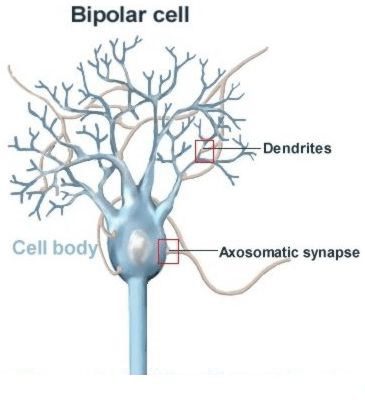
نورون ها هزاران ورودی سیناپسی دریافت می کنند که بعضی از آنها تحریکی ٬ بعضی مهاری و برخی دیگر مختلط اند. ارتباطات سیناپسی تحریکی به طور عمده در بزرگترین بخش دریافت کننده نورون ٬ دندریت ٬ یافت می شوند ؛ به ویژه روی زوائدی که از دندریت برجسته می شوند. سیناپس های تحریکی ویژگی های ریخت شناسی مشخص دارند. این سیناپس ها ٬ سیناپس های نوع Ι هستند. در بسیاری از مواقع ناحیه ای مشخص در پایانه پیش سیناپسیِ سیناپس های نوع Ι وجود دارد که مسئول آزادسازی وزیکول های حاوی گلوتامات می باشد. ناحیه ای مشابه نیز در غشای پس سیناپسی یافت می شود که برای اتصال رسپتور به گلوتامات کاربرد دارد. علاوه بر اینها ٬ وزیکول های حاوی گلوتامات بسیار کوچک هستند ( با قطری حدود ۵۰ نانومتر) و ظاهری کروی دارند.
سیناپس های مهاری (مثل سیناپس هایی که از گلیسین یا گابا به عنوان نوروترانسمیتر استفاده می کنند) بیشتر در نزدیک جسم سلولی (soma) تشکیل می شوند و به عنوان سیناپس های نوع ΙΙ شناخته می شوند. از لحاظ مورفولوژی٬ مثل سیناپس های نوع Ι ٬ نواحی اختصاصی برای آزادسازی وزیکول ها و اتصال به رسپتور وجود دارد ؛ اما این نواحی در سیناپس های نوع ΙΙ کوچکتر از سیناپس های تحریکی هستند. به دلایل نامشخص ٬ وزیکول های حاوی گلیسین یا گابا ٬ بیضی شکل هستند.
از لحاظ عملکردی، موقعیت این ارتباطات سیناپسی تاثیر بسیار زیادی روی نورون پس سیناپسی دارد. هر چقدر سیناپس از جسم سلولی دور تر باشد، پتانسیل تحریکی پس سیناپسی بیشتر توسط ویژگی های غیرفعال غشایی تضعیف می شوند (این پتانسیل ها ،پتانسیل عمل را منتشر نمی کنند ؛ بلکه پتانسیل های سیناپسی هستند). بنابراین برای نورون هایی که فاقد فرایند های رژنراتیو در دندریت هایشان هستند، EPSP هایی که دورتر از نقطه تولید پتانسیل عمل (جسم سلولی و برآمدگی آکسونی(axon hillock) ) هستند بیشتر از IPSP هایی که نزدیک تر به جسم سلولی تولید می شوند ، تضعیف می شوند. براساس این آرایش فضایی و اندازه نسبتا کوچک هر EPSP (1mv) ، تعداد زیادی EPSP در فاصله دور باید با هم جمع شوند تا باعث شروع یک پتانسیل عمل گردند. در مقایسه IPSP های موضعی کمتری روی جسم سلولی مورد نیاز هستند تا تولید پتانسیل عمل را مهار کنند. در یک نورون قشری تیپیک ممکن است ۱۰ هزار سیناپس تحریکی axodendritic یافت شود در حالی که تعداد سیناپس های مهاری axosomatic بین ۱۰-۵۰ باشد.
ساختار نوروترانسمیتر های آمینواسیدی
در ابتدا، آمینواسید ها کاندید مناسبی برای قرار گرفتن در گروه نوروترانسمیتر ها نبودند؛ زیرا اجزای سلولی فراوانی بودند که برای سنتز پروتئین ضروری بود. علاوه براین آنزیم های اختصاصی نورون ها استیل کولین و کاتکول آمین ها را می سازد اما آنزیم های سازنده گلوتامات،آسپارتات و گلیسین برای نورون ها اختصاصی نبودند. با استفاده از آنتی بادی هایی برای کولین استیل ترانسفراز می توان نورون ها را به عنوان نورون های کولینرژیک شناسایی کرد اما چنین مارکر و علامتی برای شناسایی نورون هایی که از آمینواسید به عنوان انتقال دهنده استفاده می کنند، وجود ندارد. با این حال، در حال حاضر می دانیم که اسیدهای آمینه گروه اصلی مواد مورد استفاده برای تولید پتانسیل های تحریک کننده و مهار کننده سیناپسی در CNS هستند. آمینواسیدهایی که برای انتقال سیناپسی مورد استفاده قرار می گیرند،کمپارتمان بندی شده اند. به عنوان مثال برای اینکه گلوتامات به عنوان نوروترانسمیتر مورد استفاده قرار بگیرد ، گلوتامات متابولیکی که برای سنتز پروتئین استفاده می شود توسط بسته بندی این ترانسمیتر درون وزیکول های سیناپسی کمپارتمان بندی می شود تا متعاقبا آزادسازی وابسته به کلسیم صورت گیرد.
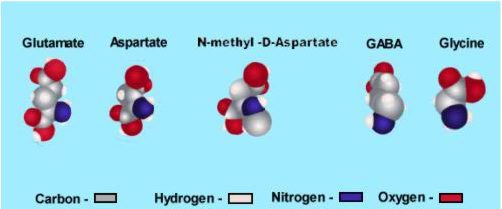
شکل بالا ساختار ۴ نوروترانسمیتر آمینواسیدی اساسی را نشان می دهد. نوروترنسمیتر های تحریکی حامل ۲ بار منفی هستند( به خاطر وجود دو گروه کربوکسیل؛ COO- ، گوی های قرمز) در حالی که آمینواسید های مهاری یک بار منفی دارند. توجه داشته باشید که N-متیل-D-آسپارتات یک ترکیب سننتیک است و در مغز یافت نمی شود؛ بنابراین به لحاظ تکنیکی یک نوروترانسمیتر نیست. این ماده یک آگونیست بسیار کاربردی است که می تواند اعمال گلوتامات را روی زیرمجموعه های خاصی از رسپتور گلوتامات تقلید کند.