مقدمه
فرض کنید که شما در حال تماشای یک مسابقه هستید؛ ضربان قلبتان از هیجان با نزدیک شدن دوندگان به خط پایان بالاتر میرود. یا فردی را میبینید که غذای ناآشنایی را بو میکشد و چهرهاش از انزجار در هم کشیده میشود، ناگهان معده شما به فکر غذا میافتد.
تا سالها چنین تجربیاتی روان شناسان، دانشمندان علوم اعصاب و فیلسوفان را متعجب کرده بود، که چرا ما چنین واکنشهای احساسی به اعمال دیگران نشان میدهیم؟ چگونه فوراً و به صورت غریزی میتوانیم افکار، احساسات و مقصود دیگران را بفهمیم؟
به باور عصب شناس ایتالیایی جاکومو ریتزولاتی (Giacomo Rizzolatti) مطالعه نورونهای عصبی میتواند در توضیح این که چگونه ذهن دیگران را میخوانیم و با آنها احساس همدلی میکنیم به ما کمک کند.
جاکومو ریتزولاتی
جاکومو ریتزولاتی متولد ۲۸ آوریل ۱۹۳۷ در کییف (Kiev)، اوکراین است. او مدرک پزشکی و تخصص علوم اعصاب خود را از دانشگاه Padua، در ایتالیا دریافت کرد. در سال ۲۰۰۲ او مدیر دپارتمان علوم اعصاب دانشگاه پارما (Parma) شد. تمرکز اصلی تحقیقات دکتر ریتزولاتی، سیستم حرکتی و نقش آن در عملکرد شناختی است. اخیراً او به مطالعه سیستم نورونهای آینهای با استفاده از تکنیکهای تصویربرداری مغز در انسان و ثبت فعالیتهای نورونی مجزا در پریماتهای غیر انسانی میپردازد.

جیاکومو ریتزولاتی
مفهوم نورون آینهای
نورونهای آینه (MNs) مجموعهای از نورونهای پیش حرکتی هستند که هم در طول فعالیت حرکتی و هم هنگام مشاهده شخص دیگری که عمل مشابه را انجام میدهد، برانگیخته میشوند. مغز انسان نورونهای بی شمار و ارتباطات عصبیای را شامل میشود که هر کدام از نظر محل و عملکرد، در این شبکه متمایز هستند.
ریتزولاتی و همکارانش به صورت تصادفی نورونهایی را در مغز میمون ماکاک (macaque) پیدا کردند که ویژگیهای منحصر به فردی داشتند؛ به این صورت که این نورونها نه تنها با فعالیت خود میمون بلکه با مشاهده انجام همان کار توسط شخص دیگر برانگیخته میشدند. این نورونها، نورونهای آینهای نامیده میشوند زیرا قادرند تا رفتار مشاهده شده را منعکس سازند و باید از سایر نورونهای قشری موجود در آن ناحیه تفکیک داده شوند. به خصوص این نورونها زمانی که فرد مشاهده کننده خود، عمل مشاهده شده را انجام داده باشد تحریک میشوند.
نورونهای آینهای عمدتا در ناحیه F5 قشر پیش حرکتی مغز ماکاک قرار دارند. در ابتدا تمرکزاین تحقیقات بر روی حیوانات بود، به عنوان مثال در پی همین پژوهشها کشف شد که نورونهای آینهای میمون نقش مهمی در عمل درک و فهم دارند. یافتههای تحقیقات حیوانی مبنای تحقیق این نورونها در مغز انسان است. در این میان نظریات گوناگونی مطرح شده است. یافتههایی وجود دارد نشان گر این که نورونهای آینهای در انسان در تکامل زبان، تقلید، همدلی و نظریه ذهن نقش دارند. با این حال، بعضی تحقیقات دیگر نشان میدهند که این نورونها تنها یک واکنش ساده به محرکهای مشاهده شده بدون هیچ گونه عملکرد درکی هستند. برخی تحقیقات دیگر حتی وجود چنین نورونهایی را در انسان نفی میکنند.
با توجه به مقاله سال ۱۹۹۶ ریتزولاتی، مطالعات انجام شده بر روی پریماتها و انسانها از این ایده که نورونهای آینهای به ما در درک رفتار مشاهده شده کمک میکنند، حمایت میکند. یک پژوهش بر روی پریماتها نشان داد که نورونهای آینهای به سادگی با صدای انجام یک عمل، مانند پاره شدن کاغذ فعال میشوند، در حالی که بر اساس پژوهش دیگر تنها تجسم ذهنی یک عمل کافی بود تا این نورونها فعال شوند. این نتایج از این نظر اهمیت دارند که نشان میدهند پاسخ نورونهای آینهای تنها به مشاهده یک عمل نیست بلکه پاسخ به مفهوم آن عمل نیز میباشد.
همچنین اگر مشاهده یک عمل و انجام همان کار قسمت یکسانی از مغز و حتی یک نورون را تحریک کند، پس میشود این استدلال را کرد که دیدن یک عمل و انجام آن هر دو میتوانند احساس یکسانی را در فرد برانگیزند. در واقع علت تکامل این سیستم در بدن تسربع فرآیند یادگیری از دیگران عنوان شده است.
یک کشف تصادفی
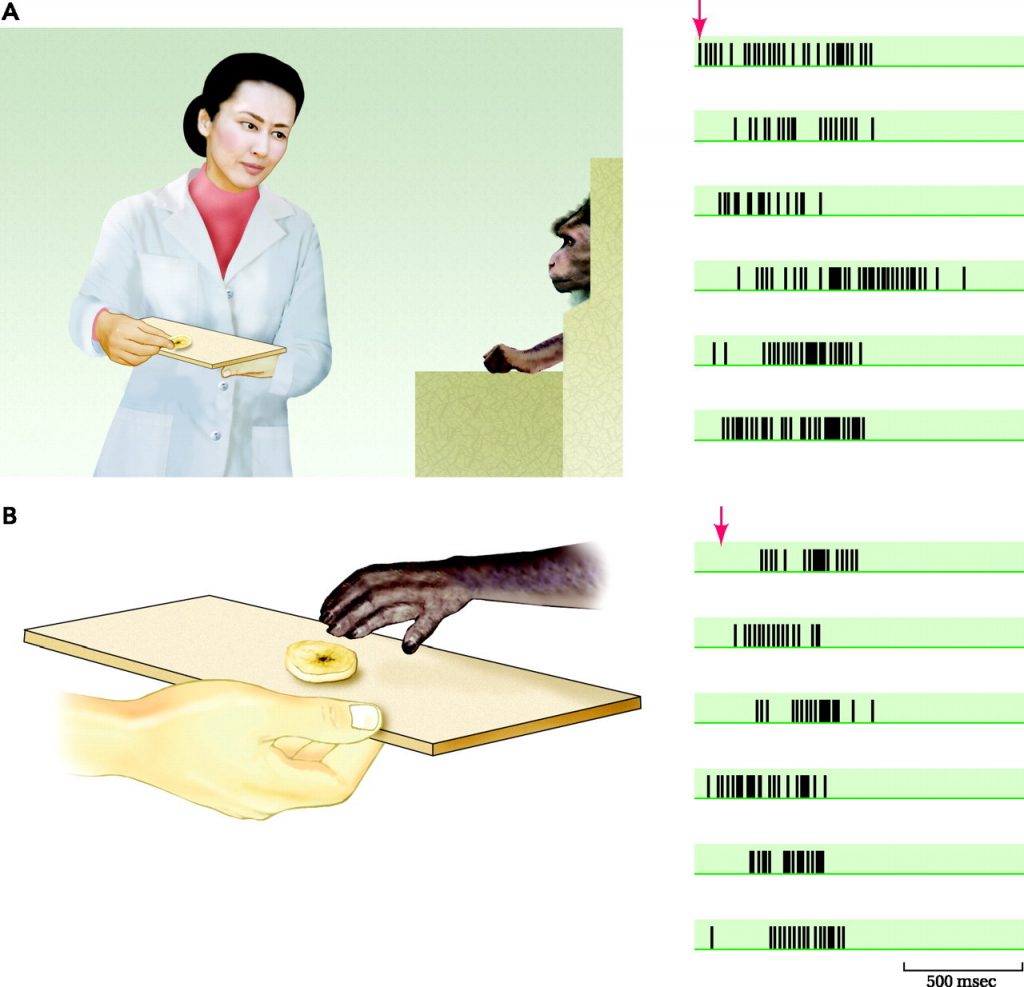
داستان کشف نورونهای آینهای بسیار ساده است. در سال ۱۹۹۲ گروهی از دانشمندان علوم اعصاب به رهبری جاکومو ریتزولاتی، الکترودهای کوچک را در مغز میمونهای ماکاک جای گذاری کردند تا بتوانند به خوبی نحوه موزون سازی تعاملات ظریف ماهیچههای دست را توسط مغز درک کنند. با استفاده از این الکترودها، محققین فعالیت نورونها را دنبال کردند. نورونها به نوعی درحکم میکروتراشههای کامپیوتری بودند. هر نورون میتواند در انجام هر کاری از تشخیص چهره گرفته تا تنظیم خواب و بیداری تخصص داشته باشد.
تیم ریتزولاتی با بررسی نورونهای مغز ماکاک، که در کنترل عضلات دست نقش داشتند، در پی کشف سلولهایی بودند که به طور ویژه در انجام عملی مانند دراز کردن دست یا در دست گرفتن شی تخصص داشته باشند. آنها نهایتاً نورونهایی را که مسئول انجام چنین کارهایی بودند را کشف کردند اما در ادامه معلوم شد که این تنها نیمی از ماجرا بوده است. یک روز هنگامی که افراد تیم در همان اتاقی که میمون نگهداری میشد غذا میخوردند، اتفاقی کاملاً غیر منتظره را مشاهده کردند: بعضی از این نورونها هنگامی فعال شدند که میمون یکی از آزمایشگران را در حال انجام کار مشابه (در این مورد، بردن غذا به سوی دهان) میدید. به طور خلاصه: این نورونها هم هنگام مشاهده و هم هنگام انجام کاری بخصوص تحریک میشدند.
به عبارت دیگر، نورونها عمل مشاهده شده را منعکس میکردند. وقتی میمون انجام یک عمل را مشاهده میکرد، مغز شبیه سازی فعالیتهای عصبی خاصی را آغاز مینمود، به گونهای که گویی میمون خود این عمل را انجام میدهد. فرض بر این است که این نورونهای آینهای سیستمی را در مغز تشکیل دادهاند که مسئول توانایی ما در درک فعالیتهای دیگران است. ما در مورد دنیای پیرامون خود میدانیم زیرا با آن تعامل داشتهایم و تنها با شبیه سازی این تعامل در مغز میتوانیم رفتار شخص دیگری را درک کنیم.
یافتههای حیوانی
پس از اولین آزمایش، تحقیقات زیادی برای پیدا کردن شواهدی در رابطه با محل و ویژگیهای نورونهای آینهای انجام شد. نتیجه این تحقیقات نشان میدهد که مناطق دیگری نیز در مغز خاصیت آینهای دارند. علاوه بر ناحیه پیش حرکتی، یکی از مهمترین نواحی ورودی، منطقه PF، ناحیه سری-پایینی لوبول آهیانهای (rostral inferior parietal lobule (IPL یا IPL)، نیز دارای نورونهایی با خصوصیات آینهای میباشد. منطقه پیش حرکتی و PF با هم ارتباط نزدیکی دارند. بیشتر نورونهای PF (حدوداً ۹۰%) به محرکهای بصری پاسخ میدهند، اما تنها نیمی از این نورونها در فعالیتهای حرکتی فعال هستند. PF خروجیهایی را به ناحیه F5 فرستاده و ورودیهایی از شکنج گیجگاهی فوقانی (Superior temporal sulcus یا STS) دریافت میکند. STS از نورونهای بصریای تشکیل شده است که به اعمال بیولوژیکی مشاهده شده مانند حرکت چهره یا بدن واکنش نشان میدهند. علاوه بر این، STS نقش مهمی در ادراک نگاه خیره دارد و بنابراین در شناخت جهت توجه دیگران نقش مهمی ایفا میکند. اگر چه STS ویژگیهای حرکتی ندارد و به عنوان بخشی از نورونهای آینهای محسوب نمیشود، اما به دلیل ارتباط نزدیکش با بخشهای تحتاتی لوب آهیانهای سیستم نورونهای آینهای، به شدت در عملکرد این سیستم تأثیرگذار است. به عنوان جمع بندی میتوان نتیجه گرفت که سیستم آینهای قشری در میمونها، منطقه شکمی پیش حرکتی و بخش سری تحتاتی لوب آهیانهای را شامل میشود.
تحقیقات اخیر نشان داده است که برخی از نورونها تنها با مشاهده فعالیتهای بسیار خاص حرکتی فعال میشوند، در حالی که دیگر نورونها با مشاهده فعالیتی که هدف مشابهی را دنبال میکنند نیز تحریک میشوند. در واقع به نظر میرسد که انواع مختلفی از نورونهای آینهای وجود دارد: strictly congruent MNs (مورد اول) وbroadly congruent MNs (مورد دوم). نورونهای آینهای از نوع broadly congruent تقریباً دو سوم جمعیت نورونهای آینهای را تشکیل میدهند.
عملکرد نورونهای آینهای در حیوانات
یکی از فرضیههای عملکرد نورونهای آینهای حیوانات، کمک به درک هدف عمل حرکتی مشاهده شده است. پژوهشگران متوجه شدند که تحریک نورونهای آینهای، به جسم درگیر در عمل یا یک ژست حرکتی خاص مربوط نیست، بلکه به معنا یا هدف عمل بستگی دارد. بنابراین پژوهشگران بر این باورند که MNها برای شناخت خودکار رفتار هدفمند اهمیت دارند.
مطالعات، علاوه بر نورونهای آینهای مبتنی بر بینایی، نشان دهنده وجود MNهای سمعی-بصری در منطقه پیش حرکتی میمونهاست. این نوع نورونها هنگامی که حیوان یک عمل را انجام میدهد و زمانی که صدای مربوط به آن عمل خاص را میشنود، توانایی تحریک شدن را دارند.
تحقیقات بیشتری در مورد ناحیه F5 قشر پیش حرکتی میمون، به طور دقیقتر قشر پیش حرکتی شکمی جانبی، نشان داده است که نورونهای آینهای واقع در آن ناحیه در طول مشاهده کارهایی که به وسیله ابزارها انجام میگردند، فعال میشوند. به اصطلاح این سلولها نورونهای آینهای tool-responding نامیده میشوند. این نورونها هم هنگام حرکات دست یا دهان میمون، هم زمانی که محققان با یک ابزار به انجام کاری مشغول میشوند، تحریک میشوند.
از دیگر یافتههای جالب این است که یک سوم از جمعیت نورونهای آینهای منطقه F5 هنگامی که میمون حرکات دهان را میبیند، فعال میشوند. فعالیت این نورونهای دهانی-آینهای (mouth mirror neurons) هنگامی رخ میدهد که حرکات مشاهده شده مربوط به عملکردهای بلعی باشد، اما در طول حرکات و ژستهای ارتباطی مانند ملچ ملوچ لبها بیشتر تحریک میشوند. این قضیه میتواند بیان گر اهمیت نورونهای آینهای در مهارتهای ارتباطی باشد. نکته جالب توجه این است که در انسان، ناحیه بروکا (Broca)، که منطقه مرتبط با مهارتهای زبانی و گفتاری است، همتای ناحیه F5 است. این امر میتواند دلالت بر نحوه تکامل زبان و گفتار در انسانها باشد.
در مطالعه دیگری با استفاده از تکنیک ثبت فعالیت یک سلول، محققین فعالیت نورونهای F5 را زمانی که آن عمل در فاصلههای مختلف از میمون انجام میشد، اندازه گیری کردند. آنها پی بردند که ۲۶٪ از نورونهای پیش حرکتی هنگامی که حرکت مشاهده شده در فضای خارج از محیط اطراف میمون، ۲۷٪ زمانی که در فضایی اطراف میمون، و ۴۷٪ بدون توجه به موقعیت عمل انجام شده تحریک میشدند. این مورد نشان میدهد که فعالیتهای مشاهده شده در فواصل مختلف از میمون به طور متفاوتی توسط نورونهای آینهای رمزگذاری میشوند. این یافتهها به طور بالقوه دلایل قویای برای رفتارهای اجتماعی و انسانی فراهم میکنند، زیرا در محیط اجتماعی مهم است که به سرعت، به رفتار مشاهده شده بر اساس مکان آن نسبت به ناظر پاسخ داده شود.
مطالعات انسانی
در انسان، دو سیستم عصبی آینهای اصلی توسط مطالعات تصویربرداری مغز شناسایی شده است. اولين سیستم واقع در لوب آهیانه، قشر پيش حرکتی و pars opecularis میباشد. pars opecularis، که بخشی از ناحیه بروکا نیز میباشد، شامل IPL و بخش دمی گیروس پیشانی تحتانی است (IFG) و یک منطقه حیاتی برای زبان و گفتار محسوب میشود. شبکه آهیانهای-پیشانی مسئول تشخیص رفتار داوطلبانه است. از آن جایی که نورونهای آینهای عمدتاً در قشر حرکتی قرار دارند، شگفت آور نیست که درست مانند این مناطق حرکتی، نورونهای آینهای یک سازماندهی سوماتوتروپیک داشته باشند. سیستم دوم آینهای شامل لوب اینسولا و قشر میانی قدامی است. سیستم آینهای لیمبیک، همانطور که از نامش پیداست، مسئول شناخت رفتار عاطفی است.
البته علاوه بر نواحی گفته شده نورونهای آینهای در نقاط دیگری از مغز نیز پراکنده شدهاند.
عملکردهای نورونهای آینهای در انسانها
اولین مطالعهای که وجود نورونهای آینهای را در مغز انسان گزارش کرد، مطالعه TMS بود.
Transcranial Magnetic Stimulation یا TMS یک روش غیر تهاجمی است که به صورت گسترده برای مطالعه نورونهای آینهای در انسان استفاده میشود. دستگاه TMS یک میدان الکترومغناطیس قوی که قادر به دپولاریزاسیون نورونهای سطحی است، را ایجاد میکند و بدین ترتیب این نورونها شروع به تولید پتانسیل عمل میکنند. مطالعات نشان دادهاند که با اعمال پالسهای تکی یا جفتی TMS به قشر حرکتی مغز انسان، یک افزایش تحریک پذیری حرکتی موقت در قشر حرکتی پدید میآید. این افزایش تحریک پذیری یک فعالیت دگرسو در ماهیچهها پدید میآورد که پتانسل برانگیخته حرکتی (Motor Evoked Potential یا MEP) نامیده میشود و به وسیله الکترومیوگرافی (EMG) از این ماهیچهها قابل اندازه گیری است.
پژوهشها نشان میدهند زمانی که ما یک عمل خاصی را مشاهده میکنیم یک افزایش اندک و زیر آستانهای در ماهیچههایی که در آن عمل شرکت میکنند رخ میدهد که به احتمال زیاد توسط نورونهای آینهای ایجاد میشود. از آن جایی که این فعالیت اندک ماهیچهای میتواند منجر به MEP شود، TMS روشی مفید برای مطالعه نورونهای آینهای میباشد.
زمانی که شرکت کنندگان آزمایشگران را در حال انجام یک سری حرکات با دستها میدیدند، با اعمال TMS بر روی قشر حرکتی، یک افزایش MEP مشاهده میشد. از آن زمان تاکنون TMS شواهد فراوانی دال بر وجود نورونهای آینهای در انسانها ارائه کرده است.
تحقیقات از این مفهوم که نورونهای آینهای انسان نیز قادر به رمزگذاری اهداف یک عمل هستند پشتیبانی میکنند. گروهی از دانشمندان طی یک مطالعه fMRI نشان دادند که اقدامات حرکتیای که جزو مجموعه اعمال یک فرد هستند (به عنوان مثال گاز گرفتن یک شی توسط سگ یا میمون) لوب آهیانه تحتانی و ناحیه pars opecularis آن را تحریک میکنند؛ بدون توجه این که این عمل توسط یک حیوان انجام شده باشد. اما زمانی که این عمل جزو رفتارهای معمول انسانی نباشد (مانند پارس کردن) تنها مناطق بصری فعال میشدند. این تجربه نشان میدهد که نورونهای آینهای انسان هنگام مشاهده عملی که جزو رفتارهای انسانیست، بدون توجه به این که انجام دهنده این عمل چه کسی است، فعال میشوند. در واقع این آزمایش نشان میدهد که MNهای انسانی با توجه به قصد و هدف یک عمل فعال میشوند، نه فقط با مشاهده ساده خود عمل.
محدوده دیگری از MNهای انسانی که توجه زیادی را جلب کرده است، گیروس لوب پیشانی تحتانی میباشد که در برگیرنده ناحیه بروکاست. پژوهشها این منطقه را با درک اجتماعی، مشاهده عمل و تقلید مرتبط میدانند.
در سال ۲۰۰۳ با استفاده از روش TMS تکراری (repetitive TMS) برای مهار منطقه، به بررسی این مورد پرداخته شد که چگونه IFG در فرایند تقلید نقش دارد. در هر آزمایش دو مجموعه پنج پالسی با سرعت ۵Hz به چپ و راست pars opercularis بخش IFG اعمال شد. نتایج نشان داد که مهار این مناطق توسط روش TMS تکراری، باعث کاهش عملکرد در وظایف تقلیدی نسبت به وظایف غیر تقلیدی، در حالی که با مهار مناطق دیگر(به طور مثال ناحیه پس سری) چنین چیزی مشاهده نشد. این آزمایش اهمیت MNS، به ویژه IFG را در رفتار تقلید نشان داد. پژوهشگران همچنین نتیجه گرفتند که منطقه بروکا نیز در تقلید نقش دارد، که خود بخشی از IFG میباشد.
این شواهد که حاکی از نقش مرکزی نورونهای آینهای در تقلید است به همراه این شواهد که نورونهای آینهای با ناحیه بروکا هم پوشانی دارد نشان میدهند که نورونهای آینهای در انسان در تکامل زبان دخیل هستند. ریتزولاتی و همکارانش (۲۰۰۴) نظریهای در این باره ارائه دادند. آنها معتقد بودند که ارتباطات مرتبط با ژستهای آوایی در بردارنده یک مفهوم هستند؛ که در ابتدا به صداهای ساده و سپس با استفاده از قابلیت تقلید نورونهای آینهای به گفتارهای پیچیده تبدیل شدهاند. همچنین وجود MN هایی که در واکنش به محرکهای معنادار شنوایی در میمونها فعال میشوند اثبات شده است.
برای ارزیابی اینکه نورونهای آینهای انسان میتوانند توسط محرکهای شنوایی فعال شود یا نه محققان به انجام یک مطالعه fMRI پرداختند. در این آزمایش، به افراد یاد داده شد که یک قطعه جدیدی را روی پیانو بنوازند، که بعداً این نت برای آنها پخش شد. در طول پروسه نواختن و گوش دادن، دادههای fMRI گردآوری شد. آنها متوجه شدند که شبکه دو طرفه پیشانی-آهیانه مرتبط با بخش حرکتی، از جمله منطقه بروکا، در طول درک شنوایی از نظر قطعه تازه و در طول اجرای آن نت فعالیت داشتهاند. این سیستم، که نویسندگان مطالعه از آن به عنوان سیستم شنوایی-انجامی (حرکتی) نام میبرند، به رپرتوار (repertoire) حرکتی فرد بستگی دارد. همان طور که به وسیله فعالیت ناشی از گوش دادن به قطعه تازه آموخته شده نشان داده شد، اعمال حرکتی تازه یاد گرفته شده میتوانند در این شبکه بسیار سریع ایجاد شوند. علاوه بر این، این سیستم به عنوان هسته مرکزی، منطقه بروکا را در اختیار دارد. به همین دلیل نویسندگان تأکید دارند که این سیستم ممکن است پیامدهای مهمی در کسب مهارت زبانی داشته باشد زیرا این سیستم شنوایی-حرکتی در طول حرف زدن بازخورد حسی-حرکتی مهمی دارد.
در نهایت با وجود اینکه محققان شواهد زیادی مبنی بر وجود نورونهای آینهای در انسانها یافتهاند اما هنوز راه زیادی در پیش روی آنهاست.