کروموزوم جنسی
اسپرم و تخمک از طریق نوعی تقسیم سلولی خاص به نام میوز تولید می شود، که تضمین می کند سلول ها فقط نیمی از کروموزوم ها را دارند – یک نسخه از هر کروموزوم به جای دو نسخه در پستانداران.
طی میوز، درهر مجموعه از کروموزوم های والدین، توالی های DNA را در مناطق معادل (هومولوگ) در طول خود رد و بدل می کنند. این مبادله ی اطلاعات – فرایندی به نام نوترکیب – ناشی از وقفه های دو رشته DNA است که تقریباً در هر ۱۰ مگابیس رخ می دهد. با این حال کروموزوم های X و Y فقط یک منطقه همولوگ کوتاه (حدود ۷۰۰ کیلوبیس) به نام منطقه شبه اتوزوم (PAR) دارند. از این رو جداشدن دو رشته باید بیشتر در این منطقه اتفاق بیفتد؛ تا از نوترکیب مناسب بین کروموزوم های X و Y اطمینان حاصل شود. Acquaviva و همکارانش در Nature درباره ی چگونگی فراهم آوردن موقعیت عنصر DNA برای افزایش دفعات شکست های دو رشته ای در PAR گزارش می دهند.
طی میوز چه اتفاقاتی می افتد؟
قسمت اولیه میوز شامل چند مرحله است. هر کروموزوم از دو نسخه یکسان که معروف به کروماتیدهای خواهری تشکیل یافته است، که توسط اجتماعی از پروتئین کوهزین به یکدیگر متصل می شوند. جداشدن دو رشته به کمک عوامل جانبی آن، توسط پروتئین SPO11 در ط.ل کروماتیدها اتفاق می افتد. انتهای ‘۳ رشته به جست وجوی توالی های همولوگ می پردازد – این کارمنجر به جفت شدن و تراز شدن کروموزوم های همولوگ (مانند X و Y یا هر دو نسخه از هر کروموزوم غیرجنسی) در طول آن ها می شود، که به آن فرآیند سیناپس گویند.
سرانجام در زمان نوترکیب، بازوهای کروماتید بین دو کروموزوم همولوگ رد و بدل می شود. این مرحله آخر برای تفکیک دقیق هر جفت کروموزوم همولوگ در سلول های دختری به طور جداگانه ضروری است. نوترکیب کافی در PAR عامل اصلی ناتوانی رشد و ناباروری است. در واقع، کروموزوم های جنسی اغلب کروموزوم های جدا از هم تفکیک شده در اسپرم هستند.
ساختمان ساقه-حلقه
فراوانی جدایی دو رشته میوتیک با تعداد سازه های کروماتید موسوم به “واحد ساقه(محور)- حلقه” ارتباط دارد.
حلقه های DNA از محور DNA غنی از پروتئین تشکیل می شوند که پایه یا اساس یا به عبارت دیگر ستون فقرات کروماتید ‘۵ را تشکیل می دهد (شکل ۱). مناطق کروماتید که دارای محورهای طولانی تر و حلقه های کوتاه تر هستند، به طور کلی دارای شکستگی های دو رشته ای نسبت به مناطق دارای محورهای کوتاه تر و حلقه های طولانی تر هستند. PAR دارای یک محور نسبتاً طولانی برای توالی ژنومی آن است، که نشان می دهد؛ ساختار کروموزوم ممکن است فرکانس بالای از هم گسیختگی برای اطمینان از نوترکیبی PAR مورد نیازش باشد.
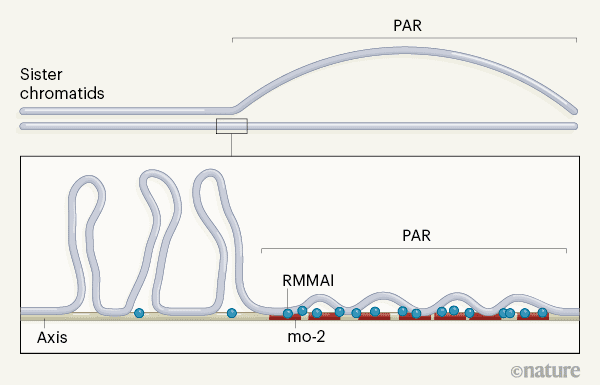
چگونه این ساختار کروموزومی خاص PAR شکل می گیرد و تنظیم می شود؟
برای پاسخ به این سوال، Acquaviva و همکاران اسپرماتوسیت های موش (پیش سازهای میوز اسپرم) تجزیه و تحلیل کردند. آن ها دریافتند که PAR قبل از سیناپسیس، برای ساختن محورهای کروماتیدهای خواهرجدا شده و برای نشکیل PAR بازسازی می شوند.(شکل ۱).
نقش و مکانیسم عملکرد پروتئین های RMMAI
نویسندگان نشان داده اند؛ که محورهای جدا شده به شدت توسط پنج پروتئین محدود می شوند که باعث ایجاد شکاف مضاعف می شود.(REC114 ،MEI4 ،MEI1 ،ANKRD31 و IHO1؛ به طور کلی RMMAI می نامند.) این فاکتورها قبلاً در نقاط دیگر هسته جمع شده بودند. Acquaviva و همکارانش دریافتند که این تجمع در مناطقی از توالی های DNA تکراری به نام min-2 minisatellites اتفاق می افتد. محققان پیشنهاد می كنند كه minisatellites mo-2 می توانند برای اتصال پروتئین ها و سایر مولكول های درگیر در بازسازی كروموزوم و ایجاد شكاف دو رشته از جمله پروتئین های RMMAI عمل كنند.
آزمایش Acquaviva و همکاران
برای آزمایش این مدل، نویسندگان یک نوع موش را مقایسه کردند که به طور طبیعی چند مینیزلات mo-2 در یک فشار آزمایشگاه استاندارد که تعداد بسیار بیشتری وجود دارد. تجمع REC114 در سویه پایین mo-2 از حیوانات بالا mo-2 کمتر بود. این گروه سپس ساختار PAR را در اسپرماتوسیت های فرزندان بین سویه های کم و زیاد-MO-2 تجزیه و تحلیل کردند. کروموزوم جنسی حاصل از والدینی که با mo-2 بالا بودند؛ همیشه سطح بالایی از غنی سازی RMMAI و بازسازی محور در PAR را نشان می دادند، صرف نظر از اینکه X یا Y بود. در مقابل، PAR ناشی از پدر و مادر کمmo-2 نیست. علاوه بر این، Acquaviva و همکارانش نشان دادند؛ که آرایش های mo-2، همراه با مقدار کافی RMMAI، می تواند به بازسازی ساختاری کروموزوم های غیرجنسی منجر شود – به عنوان مثال: در انتهای کروموزوم.
آیا تجمع RMMAI و بازسازی ساختار آرایش های mo-2 باعث افزایش فرکانس گسیختگی دو رشته می شود؟
پروتئین RPA2 در مناطقی که نوترکیبی در حال انجام است ؛ نجمع یافته اند. بنابراین، این کانون های RPA2 را به عنوان پروکسی برای شکستگی و جداشدن دو رشته مورد تحلیل قرار داد. PDK2 به شدت با آرایش های mo-2 همراه است. در واقع هنگامی که سیناپس تقریباً کامل بود، ۷۰٪ مناطق mo-2 کانون های RPA2 را درگیر کرده بودند. علاوه برطرفداری و حمایت از این ایده، این گروه نشان داد که همپوشانی بین کانون های RPA2 و آرایش های mo-2 در موش هایی که فاقد جزء RMMAI ANKRD31 بودند، به طور قابل توجهی کمتر بود و کار قبلی نشان داده است که کروموزوم های X و Y به طور صحیح در این حیوانات جفت نمی شوند.
بنابراین به نظر می رسد تجمع RMMAI برای افزایش جداشدن دو رشته و نوترکیبی در PAR مهم است. در مقابل، مکانیسم مربوط به پروتئین PRDM9 برای کنترل مناطق گسیخنگی دو رشته در خارج PAR شناخته شده است.
نویسندگان دریافتند که فرکانس جداشدن در کروموزوم های غیرجنسی در آرایش mo-2 بالاتر از حد متوسط است و نشان دادند که این استراحت ها بطور مستقل از PRDM9 شکل می گیرند. بنابراین، آرایش های mo-2 مناطق کروموزومی را تعریف می کنند که در آن مکانیسم های مستقل تشکیل PRDM9 می توانند رخ دهند، که تجمع RMMAI برای این فرایند بسیار مهم است.
جداشدن هر دو رشته و سیناپس دیرتر در آرایش mo-2 نسبت به سایر مناطق کروموزومی در اسپرماتوسیت ها اتفاق می افتد. بنابراین نویسندگان فرض کرده اند که سیناپس تاخیری ممکن است زمینه افزایش فرکانس شکاف های دو رشته درmo-2 را داشته باشد. برای آزمایش این فرضیه، آن ها ساختار کروموزوم و فرکانس جداشدن در پیش سازهای میوز تخم ها، تخمک ها را بررسی کردند. کروموزوم X تخمک دارای مناطق PAR-غنی از mo-2 است که در آن RMMAI تجمع می یابد؛ اما آن ها برای سیناپس به این مناطق متکی نیستند؛ زیرا دو کروموزوم X در طول کل مشترکات خود مشترک هستند. بنابراین کروموزوم X با همان کارایی که کروموزوم های غیرجنسی انجام می دهد سیناپس می شوند.
مدل های Acquaviva و همکاران
Acquaviva و همکاران برای نشان دادن سازگاری مدل خود که سطح شکستن دو رشته ای در PAR تخمک ها به اندازه اسپرماتوسیت ها زیاد نیست. با این حال، این گروه می تواند با تاخیر در سیناپسیس تشکیل شکاف سطح بالا در mo-2 را ایجاد کند. به طور معمول، تخمک ها با میوز زودرس بیشتر از اسپرماتوسیت ها پیشرفت می کنند. این یافته نشان می دهد؛ که مدت زمان میوز زودرس ممکن است متفاوت باشد. برای مشاهده اثرات مختلف بیولوژیکی اسپرم و تخمک تنظیم شده – در یک بازه زمانی طولانی تر مناطق آسیب پذیر مانند PAR از هم جدا گردد.
چرا محورهای کروماتیدهای خواهر در PAR از هم جدا می شوند و اینکه آیا این تفکیک باعث ایجاد شکاف و نوترکیبی می شود؟
این پدیده هنوز مشخص نشده است. به عنوان یک نتیجه از جدایی محور، کروماتیدهای خواهری کروموزوم های جنسی اسپرماتوسیتی از سایر کروموزوم ها دورتر هستند.
نویسندگان پیشنهاد می کنند که در جدایی محور ممکن است که نوعی نوترکیب داخلی غیر موثر را به نفع نوترکیب همولوگ بین کروموزوم ها سرکوب کند. علاوه بر این، این گروه بیان می کنند که تغییر ساختار کروموزوم جنسی با تجمع یک پروتئین کوزین که در نوک PAR مشاهده می شود، مرتبط است. تجمع کوزین در این منطقه ممکن است انسجام بین کروماتیدهای خواهر را افزایش دهد، که به نوبه خود به تثبیت ارتباط بین کروموزوم های همولوگ کمک می کند. یک مدل جایگزین این است که جداسازی محور ممکن است “real estate” بیشتری را فراهم کند که در آن فاکتورهای افزایش شکست های دوطرفه می توانند در PAR جمع شوند؛ زیرا پروتئین ها می توانند بین محورهای جدا شده جمع شوند. این مدل های غیر انحصاری برای تحقیقات آینده جذابیت دارند.
چگونگی شکل گیری جداشدن دو رشته در PAR، یک معضل برای این زمینه بوده است. مدل Acquaviva و همکارانش تصویری زیبا از چگونگی تعامل یک عنصر ژنتیکی، ساختار کروموزوم جنسی و زمان میوز را برای اطمینان از نوترکیب مناسب نشان می دهند.