درک مکانیسم همانندسازی دئوکسی ریبونوکلوئیک اسید (DNA) مدیون دههها تلاش دانشمندان است. به زبانی ساده، همانندسازی replication) DNA)، به استفاده از یک رشته از آن، به عنوان الگو به منظور ساختن رشتهی جدید و یکسان با رشتهی مقابل میگویند. آنزیمشناس و برندهی جایزهی نوبل اهل آمریکا، آرتور کورنبرگ، این پروسه را با ضبط نوار دستورالعمل برای اجرای یک کار مقایسه کرده است: “رونوشتهای دقیقی با این عمل بدست میآید؛ درست مثل ضبط نوار. بنابراین این اطلاعات میتواند دوباره و هرجایی در زمان و فضا مورد استفاده قرار گیرد”.
در واقعیت ولی، پروسهی همانندسازی بسیار پیچیدهتر از چیزی که کورنبرگ پیشنهاد میکند. محققان معمولاً در آزمایشهای خود از سلولهای سادهی باکتریایی استفاده میکنند اما هنوز به همهی جوابها نرسیدهاند؛ مخصوصاً وقتی بحث، همانندسازی یوکاریوتهاست. با این وجود، دانشمندان با گامهای پایهای در پروسهی همانندسازی آشنایی دارند و به همین اطلاعات اتکا میکنند تا آزمایشها و پژوهشها را به پیش ببرند.
مکانیسم مولکولی در همانندسازی DNA باکتری
یک سلول نرمال باکتریایی چیزی در حدود یک تا چهار میلیون جفت باز DNA دارد؛ این رقم را با سه میلیارد جفت باز در ژنوم موش خانگی مقایسه کنید. با این حال در همین باکتریهای ساده نیز همانندسازی DNA به طور غیرقابل تصوری پیچیده است و با یک سری وقایع مولکولی پشت سر هم ارتباط دارد. این وقایع را میتوان در چهار مرحلهی اصلی خلاصه کرد: آغاز (initiation)، باز کردن پیچخوردگی (unwinding)، سنتز پرایمر (primer synthesis) و طویلسازی (elongation).
آغاز و باز کردن پیچخوردگی
در طول مرحلهی آغاز، پروتئینهای به اصطلاح آغازگر، به محل همانندسازی (توالی نوکلئوتیدی به نام oriC) متصل میشوند. این اتصال وقایعی را رقم میزند که در نهایت به باز کردن پیچخوردگی DNA و تبدیل آن به دو مولکول DNA تکرشتهای میانجامد. چندین گروه پروتئینی در این کار دخیل اند (شکل ۱)؛ برای مثال، DNA هلیکاز مسئول باز کردن دو رشتهی مکمل با شکستن پیوندهای هیدروژنی بین بازهاست. این پیوندهای هیدروژنی مشخصههای ضروری مدل DNA سه بعدی ارائه شده توسط واتسون و کریک است. بخاطر تمایل رشتههای جدا شده برای پیوند دوباره با یکدیگر، مولکولهایی به نام پروتئینهای متصل شونده به تک رشته، این رشتههای را پایدار نگه میدارند تا طویلسازی شروع شود. خانوادهی سوم پروتئینها به نام توپوایزومرازها بخشی از کشیدگی ناشی از پیچخورگی را بر اثر جدا شدن مارپیچ دو رشتهای کاهش میدهد.
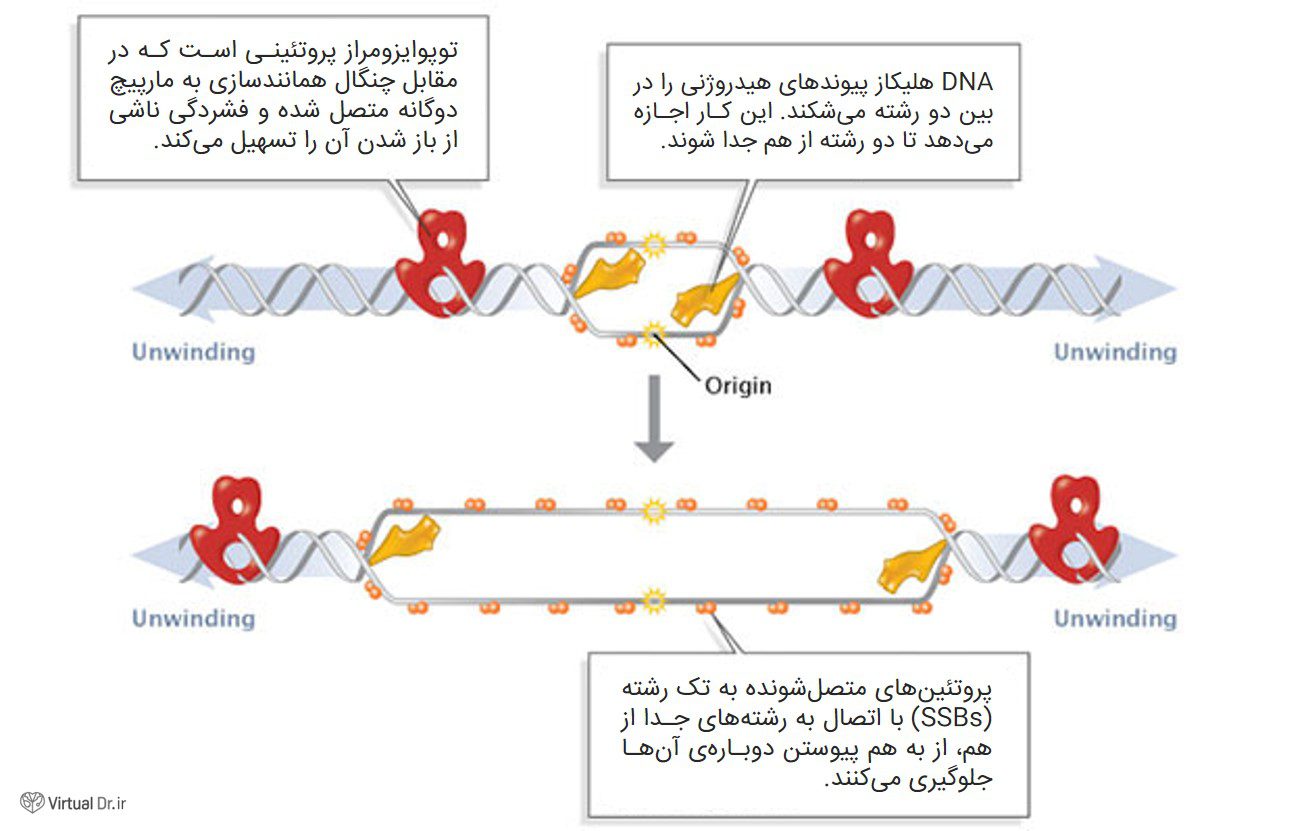
همانطور که قبلاً اشاره شد، محلی که DNA از آن جا شروع به باز شدن میکند، مبدأ (origin) همانندسازی نام دارد. همانطور که در شکل ۱ نشان داده شده، با باز شدن دو رشتهی DNA، همانندسازی به طور همزمان در دو رشته شروع میشود اما در جهتهای مخالف؛ مثلاً یکی از راست به چپ روی یک رشته و دیگری از چپ به راست در رشتهی دیگر. این باعث میشود دو چنگال همانندسازی به وجود بیاید که در طول DNA حرکت و همانندسازی میکنند.
سنتز پرایمر
این مرحله از سنتز واقعی مولکول DNA جدید شروع میشود. پرایمرها، قطعات کوتاهی از نوکلئوتیدها هستند (تقریباً ب طول ۱۰ تا ۱۲ باز) که توسط یک آنزیم RNA پلیمراز به نام پرایماز سنتز میشود. وجود پرایمرها ضروری است؛ چون DNA پلیمراز که مسئول اصلی اضافه کردن نوکلئوتیدهای جدید است، فقط میتواند دئوکسی ریبو نوکلئوتید را به گروه ۳′-OH موجود در زنجیره اضافه نماید و سنتز از نو (de novo) کار آن نیست. سنتز از نو کاریست که پرایماز آن را انجام میدهد. پس از تکمیل طویلسازی، پرایمر برداشته شده و با نوکلئوتیدهای DNA جایگزین میشود.
طویلسازی
در نهایت طویلسازی (اضافه کردن نوکلئوتیدها به رشتهی جدید DNA) پس از اضافه شدن پرایمر شروع میشود. سنتز رشتهی در حال ساخته شدن شامل اضافه کردن نوکلئوتیدها، یکی پس از دیگری، دقیقاً با ترتیبی است که بر اساس رشتهی الگوی بخصوص میباشد. حتماً هم میدانید که یکی از ویژگیهای مدل واتسون و کریک جفت شدن بازهای آدنین و تیمین با یکدیگر و گوانین و سیتوزین هم با هم است.
DNA همیشه در جهت ۵′ به ۳′ سنتز میشود؛ به این معنا که نوکلئوتیدها فقط به انتهای ۳′ رشتهی در حال رشد اضافه میشوند. همانطور که در شکل ۲ نشان داده شده، گروه ۵′-فسفات نوکلئوتید جدید به گروه ۳′-OH آخرین نوکلئونید متصل میشود. دانشمندان هنوز پلیمرازی را شناسایی نکردهاند که بتواند بازها را به انتهای ۵′ رشتهی DNA وصل کند.
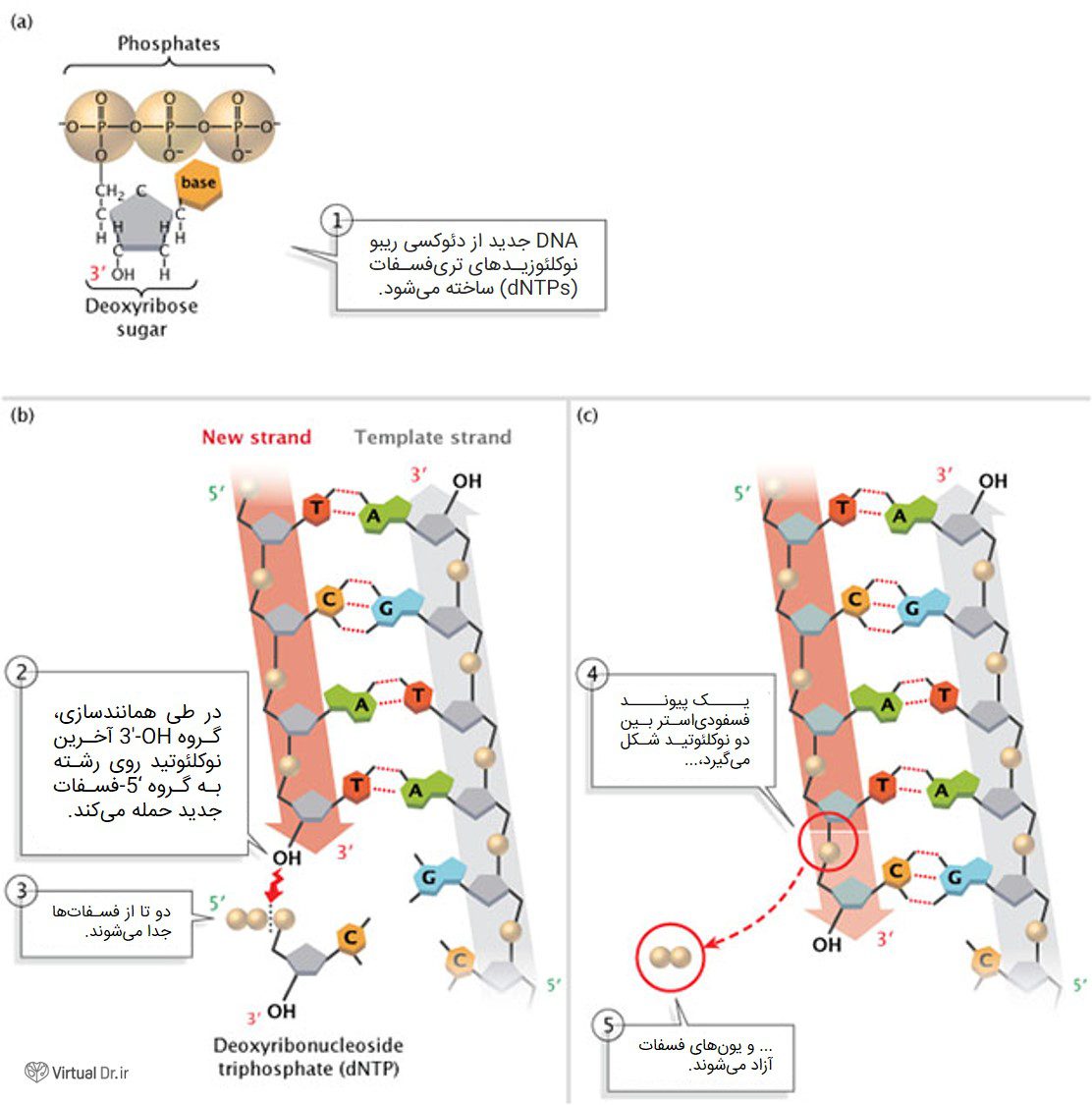
شکل ۲
کشف DNA پلیمراز
آرتور کورنبرگ در حال مطالعهی باکتری E. coli بود که متوجه شد DNA پلیمراز سنتز DNA را کاتالیز میکند. آزمایش او شامل اضافه کردن همهی مواد ضروری برای سنتز DNA باکتری E. coli در لولهی آزمایش بود که شامل نوکلئوتیدها، عصارهی باکتری و ATP میشد. او سپس خالصسازی انجام داده و آنزیمهای درگیر را تست کرد. با استفاده از این روش، کورنبرگ نه تنها DNA پلیمراز را کشف کرد، بلکه برخی کارهای اولیه را برای نمایش نقش آنزیمها در اضافه کردن نوکلئوتیدهای جدید به رشتهی DNA به انجام رساند.
دانشمندان تاکنون در مجموع پنج DNA پلیمراز متفاوت را در E. coli شناسایی کردهاند که هر کدام وظیفهی مشخصی دارد. برای مثال، DNA پلیمراز III بیشتر کار طویلسازی را با اضافه کردن یک به یک نوکلئوتیدها به انتهای ۳′ رشتهی در حال رشد انجام میدهد. بقیهی آنزیمها، شامل DNA پلیمراز I و RNase H، مسئول برداشتن پرایمرهای RNA بعد از شروع کار DNA پلیمراز III میباشد و آن را با نوکلئوتیدهای DNA جایگزین میکند. وقتی این کارها تمام شد، شکافی بین DNA حاصل از طویلسازی و DNA حاصل جایگزینی پرایمرها وجود دارد. آنزیم دیگری به نام DNA لیگاز این رشتهها را در محل نوکلئوتیدهای مجاور به هم میدوزد.
DNA پلیمراز فقط در یک جهت حرکت میکند
پس از سنتز یک پرایمر روی یکی از رشتههای DNA و جدا شدن دو رشته از هم، سنتز و طویلسازی فقط در یک جهت قابل پیشروی است. همانطور که قبلاً ذکر شد، DNA پلیمراز فقط میتواند انتهای ۳′ را طویلتر کند؛ بنابراین انتهای ۵′ پرایمر دستنخورده باقی میماند. متعاقباً سنتز فقط در رشتهای پیش میرود که رشتهی رهبر (leading strand) نام دارد. این همانندسازیِ بدون معطلی را به عنوان همانندسازی پیوسته در نظر میگیریم. رشتهی دیگر (در ۵′ از پرایمر) رشتهی پیرو (lagging strand) نام دارد و همانندسازی روی آن غیرپیوسته انجام میگیرد. مارپیچ دو رشتهای کمی قبل از این که سنتز پرایمر دیگر شروع گردد، باید از هم جدا شود. سنتز از انتهای ۳′ پرایمر جدید آغاز میگردد. پس از آن مارپیچ کمی بیشتر باز شده و جهش همانندسازی بعدی به پیش میرود. در نتیجه، همانندسازی در طول رشتهی پیرو فقط میتواند به صورت ناپیوسته انجام گیرد (شکل ۳).
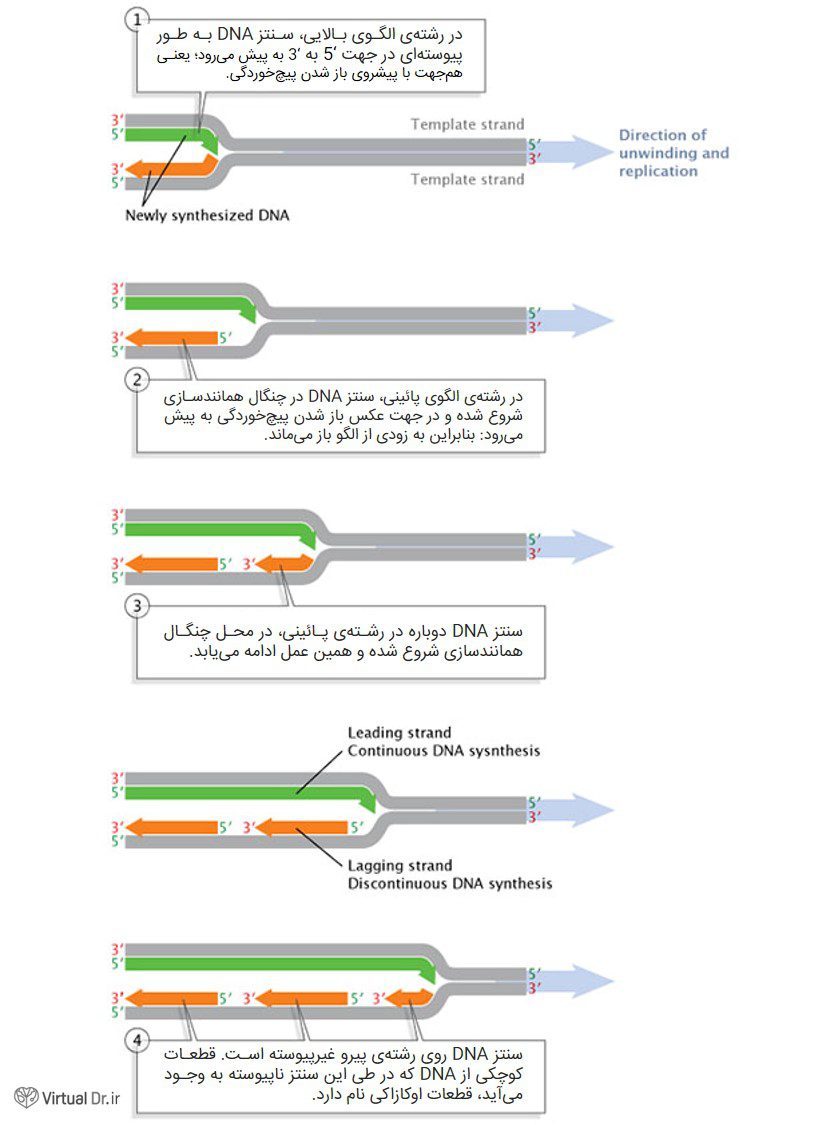
قطعات DNA تازه ساخته شده در طول رشتهی پیرو، قطعات اوکازاکی نام دارد و به اسم بیولوژیست ژاپنی و کاشف آنها، ریجی اوکازاکی، نامگذاری شده است. اوکازاکی و همکارانش با هدایت چیزی که با نام آزمایش pulse-chase میشناسیم به این کشف نائل شدند.
این آزمایش شامل قرار دادن DNA در حال همانندسازی در معرض یک پالس کوتاه از نوکلئوتیدهای نشاندار با ایزوتوپ میباشد و سپس با مدت زمانی که سلولها در معرض نوکئوتیدهای غیرنشاندار قرار میگیرند، میتوان تفاوت ایجاد کرد. این فاز ثانویه همان “chase” است. نوکلئوتیدهای نشاندار فقط در طول چند ثانیهی ابتدائیِ پالس به DNA در حال رشد میپیوندند؛ بنابراین فقط نوکلئوتیدهای غیرنشاندار در فاز chase شرکت میکنند. پس از سانتریفیوژ DNA تازه ساخته شده، مشاهده شد که فازهای ثانویهی کوتاهتر در بسیاری از فعالیتهای رادیواکتیو در DNA به اصطلاح “آهسته” دیده میشود. میزان رسوب بر اساس اندازه تعیین میشود: قطعات کوچکتر بسیار آهستهتر از قطعات بزرگتر رسوب میکنند؛ چون وزنشان کمتر است. با طولانی کردن فاز دوم، رادیواکتیویتی در DNA “سریع” افزایش مییابد؛ این در حالی است که در DNA “آهسته” اگر نگوئیم هیچ، رادیواکتیویتی کمی دیده میشود.
محققان به درستی این مشاهدات را تفسیر کردند: با فازهای کوتاهتر، فقط قطعات بسیار کوچک DNA در طول رشتهی پیرو سنتز میشود. وقتی زمان بیشتری به همانندسازی داده شود، قطعات حاصل از رشتهی پیرو، طولانیتر و سنگینتر خواهند شد؛ بنابراین سریعتر رسوب خواهند کرد. امروزه دانشمندان میدانند که قطعات اوکازاکی در DNA باکتری معمولاً طولی به اندازهی ۱,۰۰۰ تا ۲,۰۰۰ نوکلئوتید دارند؛ در حالی که در سلولهای یوکاریوتی این اندازه کوچکتر هم میباشد؛ تقریباً قطعهای به اندازهی ۱۰۰ تا ۲۰۰ نوکلئوتید.
چالشهایی که در مطالعهی همانندسازی در یوکاریوتها وجود دارد
باکتریها و یوکاریوتها در بسیاری از ویژگیهای پایهای همانندسازی اشتراک دارند؛ برای مثال، مرحلهی آغاز نیازمند پرایمر است، طویلسازی همیشه از ۵′ به ۳′ انجام میشود و همیشه در رشتهی رهبر پیوسته و در رشتهی پیرو ناپیوسته میباشد.
با این حال به تفاوتهای مهمی نیز در این باره میتوان اشاره کرد که بیولوژیستها بدون وقفه به دنبال کشف جزئیات مولکولی آنها هستند. یکی از آنها این است که همانندسازی در یوکاریوتها مبدأهای بسیاری دارد (اغلب هزاران)، نه فقط یکی و توالی این مبادی به طور گستردهای بین گونههای مختلف، متفاوت است. از طرفی دیگر، با این که مبدأهای همانندسازی در باکتریها (oriC) از نظر طول (از ۲۰۰ جفت باز گرفته تا ۱,۰۰۰) و توالی با هم متفاوت اند، به جز در بین ارگانیسمهای نزدیک به هم، با این وجود همهی باکتریها فقط یک مبدأ همانندسازی دارند.
همچنین همانندسازی در یوکاریوتها با DNA پلیمرازهای متفاوتی انجام میگیرد (مثلاً DNA پلیمراز δ و ε به جای DNA پلیمراز III). دانشمندان هنوز در حال مطالعهی نقش ۱۳ پلیمرازی هستند که تا امروز پیدا کردهاند. علاوه بر این، در یوکاریوتها، DNA الگو با پیچیدن به دور پروتئینهایی به نام هیستون فشرده میشود. این کمپلکس DNA-هیستون که نوکلئوزوم نام دارد، کار همانندسازی DNA را برای سلول و مطالعهی آن را برای دانشمندان دشوار میکند. در طول همانندسازی چه اتفاقی برای نوکلئوزوم میافتد؟ دانشمندان از مطالعهی میکروگرافهای الکترونی به این نتیجه رسیدهاند که نوکلئوزومها با فاصلهی کوتاهی پس از همانندسازی قابل مشاهده اند (نوکلئوزوم در تصاویر میکروسکوپ الکترونی قابل مشاهده است). با این حال جزئیات این اتفاقات معلوم نیست.
یادمان نرود که DNA یوکاریوت بر خلاف باکتری، خطی است. در همانندسازی DNA حلقوی، پرایمر سریعاً با نوکلئوتید تعویض میشود و خلأای را در بین DNA تازه سنتز شده باقی نمیگذارد. بر خلاف آن، در همانندسازی DNA خطی، همیشه فاصلهای در انتهای کروموزوم وجود دارد؛ چون گروه ۳′-OH برای جایگزینی موجود نیست. اگر هیچ راهی برای پر کردن این خلأ وجود نداشت، مولکول DNA از هر نسل به نسل بعدی کوتاهتر و کوتاهتر میشد. ولی این گونه نیست؛ انتهای کروموزومهای خطی، یعنی تلومرها، چندین ویژگی دارند که از این امر جلوگیری میکند.
همانندسازی DNA در فاز سنتز (S) تقسیم سلولی انجام میگیرد. این به آن معناست که در E. coli، همانندسازی کل ژنوم فقط ۴۰ دقیقه طول میکشد تا کامل شود؛ با سرعتی در حدود ۱,۰۰۰ نوکلئوتید در ثانیه. در یوکاریوتها، سرعت بسیار پائینتر است: تقریباً ۴۰ نوکلئوتید در ثانیه. هماهنگی کمپلکسهای پروتئینی برای مراحل همانندسازی و سرعتی که در آن همانندسازی باید به ترتیب تقسیم سلولی اتفاق بیافتد، بر این امر تأثیرگذارند؛ باید دقت بالای همانندسازی (proofreading) در یوکاریوتها را هم در این سرعت پائینتر دخیل دانست؛ خطاهای بسیار کمی از چشم این سیستم دور میماند.
خلاصه
مطالعهی همانندسازی DNA، تقریباً همعصر با شناسایی ساختار آن شروع شد و تا همین امروز ادامه دارد. هماکنون، مراحل آغاز، باز شدن پیچخوردگی، سنتز پرایمر و طویلسازی با پایهایترین مکانیسمهایشان شناخته شدهاند، ولی سؤالات بسیاری باقیست؛ مخصوصاً وقتی موضوع بحث، همانندسازی در یوکاریوتهاست. دانشمندان هزاران سالنفر برای مطالعهی همانندسازی وقف کردهاند. محققانی مثل کورنبرگ و اوکازاکی فقط تعدای از آنها هستند که موفقیتشان چشمگیر بوده. با این وجود، مثل بقیهی زمینههای علوم طبیعی، چیزهای زیادی برای آموختن وجود دارد؛ مثلاً این که چگونه خطاهای همانندسازی به شکلگیری بیماریهای انسانی کمک میکنند.