قبل از شروع این مطلب و بررسی پاتوفیزیولوژی بیماری پارکینسون ، خواندن این مقاله را توصیه میکنیم.
آناتومی
بیماری پارکینسون عمدتا اختلال گانگلیونهای بازال یا عقدههای قاعدهای است. گانگلیونهای بازال گروهی از هستههای واقعشده در قاعدهی مغز پیشین هستند. استریاتوم، متشکل از هسته دمدار (caudate) و هسته پوتامن (putamen)، بزرگترین مجموعهی هستهای گانگلیونهای بازال است. استریاتوم ورودیهای تحریکی را از نواحی مختلف کورتکس مغز و همچنین ورودیهای مهاری و تحریکی را از سلولهای دوپامینرژیک بخش متراکم ماده سیاه (substantia nigra pars compacta یا SNc) دریافت میکند. این ورودی ها توسط نورونهای خاردار (spiny projection neurons) دریافت میشوند که دو نوع هستند: نورونهایی که مستقیما به بخش داخلی گلوبوس پالیدوس (GPi)، خروجی اصلی گانگلیونهای بازال، و نورونهایی که به بخش خارجی گلوبوس پالیدوس (GPe) که مسیری فرعی از طریق هسته ساب تالامیک به GPi ایجاد میکند، منتهی میشوند.
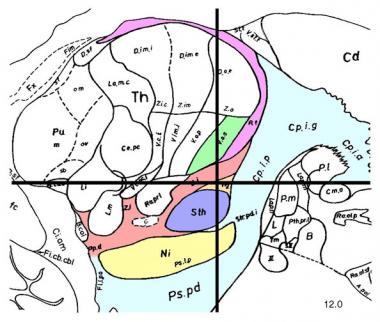
فعالیتهای مسیرهای مستقیم و غیرمستقیم، خروجیهای نورونی GPi را تنظیم میکنند که ورودیهای مهاری تونیک برای هستههای تالامیک میفرستد که به نواحی اولیه و مکمل حرکتی منتهی میشود.
پاتوفیزیولوژی بیماری پارکینسون
هیچ معیار خاص و استانداردی برای تشخیص نوروپاتولوژیک و مکانیسم بیماری پارکینسون وجود ندارد، زیرا اختصاصیت و حساسیت یافتههای مشخصه آن به روشنی مشخص نشده است. با این حال، دو یافتهی نوروپاتولوژیک اصلی در بیماری پارکینسون وجود دارد:
- از بین رفتن نورونهای دوپامینرژیک پیگمانته بخش متراکم ماده سیاه (SNc)
- وجود اجسام لویی (Lewy bodies) یا نوریتهای لویی (Lewy neurites)
از بین رفتن نورونهای دوپامین غالبا در بخش جانبی-شکمی سابستنشیا نیگرا رخ میدهد. تقریبا ۶۰ تا ۸۰ درصد نورونهای دوپامینرژیک پیش از ظهور علائم حرکتی بیماری پارکینسون از بین میروند.
در کالبدشکافی برخی از افرادی که در زمان مرگشان بهنظر میرسید از نظر نورولوژیکی نرمال هستند، اجسام لویی یافت شده است. فرض میشود که این اجسام لویی که اتفاقی کشف میشوند، نشاندهندهی مرحلهی قبل از ظهور علائم بیماری پارکینسون هستند. شیوع وجود اجسام لویی با افزایش سن بیشتر میشود. توجه داشته باشید که اجسام لویی اختصاصی بیماری پارکینسون نیست، زیرا در برخی از موارد پارکینسونیسم آتیپیک، بیماری هالرووردن ـ اسپاتز (Hallervorden-Spatz) و سایر اختلالات نیز یافت میشود. با این حال، یافتهی پاتولوژیکی تشخیصی برای بیماری پارکینسون است.
مدار حرکتی بیماری پارکینسون
مدار حرکتی گانگلیونهای بازال خروجیهای کورتکس را که برای حرکات طبیعی لازم هستند را تعدیل میکند.
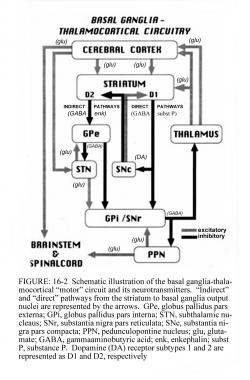
سیگنالهای حاصل از کورتکس مغز از طریق مدار حرکتی گانگلیونهای بازال-تالاموکورتیکال پردازش میشوند و از طریق مسیر فیدبک به همان ناحیه برمیگردند. خروجیهای مدار حرکتی از طریق بخش داخلی گلوبوس پالیدوس (GPi) و بخش مشبکی ماده سیاه (substantia nigra pars reticulate یا SNr) هدایت میشوند. این خروجیهای مهاری به مسیر تالاموکورتیکال هدایت شده و حرکات را سرکوب میکنند.
دو مسیر در مدار گانگلیونهای بازال وجود دارد: مسیر مستقیم و مسیر غیرمستقیم
در مسیر مستقیم، جریان خروجی از استریاتوم مستقیما GPi و SNr مهار میکند. نورونهای استریاتوم شامل رسپتورهای D1 مسیر مستقیم را تشکیل میدهند و به GPi/SNr منتهی میشوند.
مسیر غیرمستقیم شامل اتصالات مهاری بین استریاتوم و بخش خارجی گلوبوس پالیدوس (GPe) و بین GPe و هسته ساب تالامیک (STN ) است. نورونهای استریاتوم و رسپتورهای D2 بخشی از مسیر غیرمستقیم هستند و به GPe منتهی میشوند.
هسته ساب تالامیک اثر تحریکی روی GPi و SNr دارد. GPi/SNr خروجیهای مهاری را به هسته جانبی-شکمی تالاموس میفرستد. دوپامین از نورونهای نیگرواستریاتال (substantia nigra pars compacta) برای فعالسازی مسیر مستقیم و مهار مسیر غیرمستقیم آزاد میشود. در بیماری پارکینسون، کاهش دوپامین استریاتوم از طریق هردو مسیر مستقیم و غیرمستقیم منجر به افزایش خروجیهای مهاری از GPi/SNr میشود.
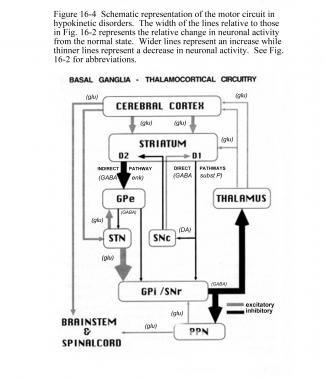
افزایش مهار مسیر کورتیکوتالامیک، حرکات را سرکوب میکند. از طریق مسیر مستقیم، کاهش تحریک دوپامین استریاتوم منجر به کاهش مهار GPi/SNr میشود. از طریق مسیر غیرمستقیم، کاهش مهار دوپامین منجر به افزایش مهار GPe و درنتیجه عدم مهار STN میشود. افزایش خروجی STN، خروجی مهاری GPi/SNr به تالاموس را افزایش میدهد.