گسترش راهی مؤثر و قابل اعتماد برای ایجاد تغییرات دقیق و هدفدار روی ژنوم سلولهای زنده، هدفی دور و دراز و باارزش در تحقیقات پزشکی و زیستشناسی محسوب میشود. اخیراً کشف آنزیم نوکلئازی به نام “پروتئین-۹ نوکلئاز مرتبط با CRISPR” یا همان Cas9 از باکتری Streptococcus pyogenes اتفاقات هیجانانگیزی را در دنیای علم رقم زدهاست.
این روش چندین هدف را در طول چندین سال برای دستکاری در ژنها دنبال کردهاست؛ از جمله کراسینگاوور بین کروموزومهای همولوگ، مداخلهی RNAها که به طور اختصاصی جزوی از اساس آزمایشگاهی به منظور ایجاد توانایی برای کسب اطلاعات عملکردی از ژن میباشد. اما این مورد با تدارک مهاری موقتی از عملکرد ژن و اثرات دور از هدف غیرقابل پیشبینی آن محدود میشود. این تکنیکها با ایجاد جهشهای دائمی به وسیلهی شکستن دو رشتهی DNA و فعال کردن مسیر ترمیم آنها موجب اصلاح جهشهای ژنتیکی میشوند ولی هیچکدام از آنها در کارایی و عملکرد در تحقیقات بزرگ مقیاس به پای روش CRISPR) Cas9) نمیرسند.
مکانیسم عملکرد سیستم CRISPR/Cas9
اول بهتر است ببینیم این اصطلاح مخفف کدام کلمات است؛ CRISPR مخفف عبارت “تکرارهای متقارن کوتاه و فاصلهدار و منظم خوشهای” میباشد و ژنهای مرتبط با آن برای ایمنی و انطباق در باکتریها و آرکیباکتریها ضروری هستند و این موجودات برای پاسخدهی و حذف ژنهای تهاجمی از آنها استفاده میکنند. این تکرارها در ابتدا در دههی ۱۹۸۰ در E.coli شناسایی شدند ولی عملکرد آنها تا سال ۲۰۰۷ مورد تأیید قرار نگرفته بود. تا این که در باکتری ترموفیلوس مشاهده شد که میتواند با برش ویروس عفونی به جایگاههای ژنی CRISPR، مقاومت بدست بیاورد.
سه نوع از مکانیسمهای CRISPR تاکنون شناسایی شده و نوع دوم آن پذیرای بیشترین مطالعات بوده است. در این نمونه DNAهای متجاوز از ویروسها و پلازمیدها به قطعات کوچکی بریده شده و جایگاههای CRISPR را شکل میدهند که در میان توالیای از تکرارها قرار میگیرد (تقریباً ۲۰ جفت باز). این قطعات سپس ترجمه شده و RNAهای کوچکی را به وجود میآورند. این مولکولها عوامل تجزیهکنندهی DNAهای متجاوز را بر اساس مکمل بودن توالیها هدایت میکنند. در واقع با الگوبرداری از توالی ژنی DNA خارجی برای فعالیت نوکلئازی سلول باکتری در برابر آنها اقدام صورت میگیرد.
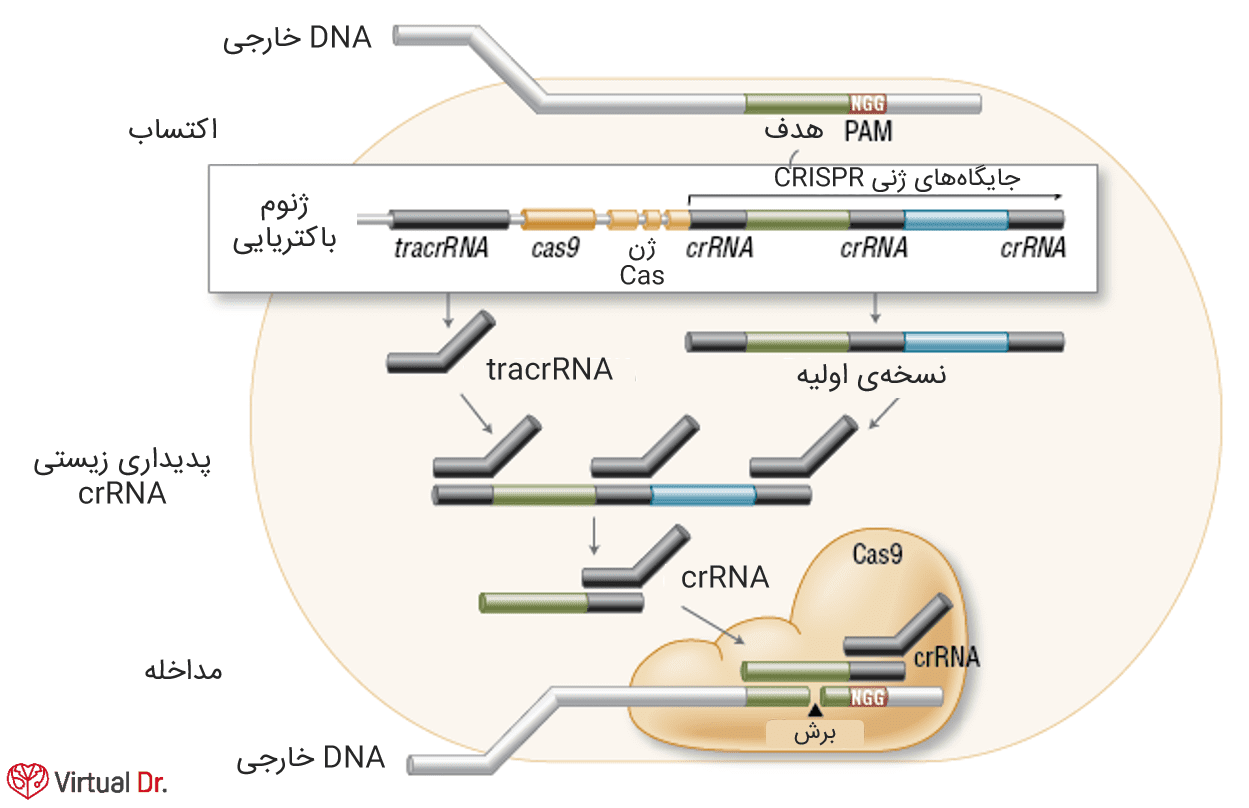
در مرحلهی اکتساب، DNA خارجی و ژنوم باکتری در جایگاههای CRISPR به هم میپیوندند. سپس این جایگاهها ترجمه شده و crRNAها را میسازند. در طی این مداخله آنزیم اندونوکلئاز Cas9 با crRNA و tracrRNA کمپلکسی را تشکیل میدهد که DNA خارجیِ حاوی ۲۰ نوکلئوتید را خواهد شکست.
پروتئینی وابسته به CRISPR به نام Cas9 به عنوان بازیگری کلیدی در مکانیسم مورد نظر نشان داده شده است. نوع دوم این مکانیسم در مقایسه با انواع دیگر منحصر به فرد است از این نظر که پروتئین Cas برای خاموشی ژن ضروری است. Cas9 در پردازش crRNAها شرکت میکند و مسئول تخریب DNA هدف است و در هر دوی این واکنشها به وجود دو دامنهی نوکلئازی وابسته میباشد. برای دستیابی به برش DNA، آنزیم مورد بحث ما باید با دو RNA هماهنگ شود؛ crRNA و tracrRNA (برای بلوغ crRNA لازم است).
در طی تخریب DNA دو دامنهی نوکلئازی نامبرده هر دو رشتهی DNA را برش داده و شکستگیهای دورشتهای را در محلی که ۲۰ نوکلئوتید دارد، همراه با رونویسی از crRNA وابسته به وجود میآورد. یکی از دامنهها رشته مکمل و دیگری غیرمکمل را ایجاد خواهدکرد.
ابزاری جدید در بیولوژی مولکولی
سادگی نوکلئازی CRISPR نوع ۲ تنها با سه جزء لازم برای انجام کل فرآیند (Cas9 همراه با crRNA و tracrRNA) این سیستم را برای اطلاح ژنی ایدئال میسازد. این پتانسیل در سال ۲۰۱۲ شناخته شد؛ پس تکنولوژی جدیدی است. بر پایهی سیستمی که در بخش قبل توضیح دادیم، سیستم سادهای دوجزئی از طریق ترکیب tracrRNA و crRNA به تک RNAای راهنما (sgRNA) به وجود آمده و به اندازهی سیستم دوجزئی قبل کارآمدی خود را در تغییر ژنها نشان داده است.
تا امروز ۳ نوع مختلف از آنزیم Cas9 برای تغییرات ژنی معتبر شناخته شده است. نوع اول که نوع وحشی آن نیز هست میتواند به طور اختصاصی دو رشتههایی از DNA جدا کند که نتیجهی آن فعال شدن ماشینهای ترمیم دورشتهایهای جدا (DSBs) است. این دو رشتهایها میتوانند توسط اتصال انتهای غیرهمولوگ سلولی ترمیم شوند. نتیجه الحاق یا حذف خواهد بود که مکان هندسی هدف را از هم خواهد گسست. متناوباً اگر تمپلیت اهدایی با همولوگی برای مکان هندسی هدف تهیه شود، دورشتهای جدا شاید با مسیر ترمیم مستقیم همولوگی (HDR) ترمیم شود که به ایجاد جایگیری جهشهای دقیق خواهد انجامید.
سیستم Cas9 قدمی دیگر برای دقیقتر شدن پیش رفته. این کار با ایجاد فرمی تغییرپذیر، تنها با فعالیت نیکازی ممکن شده است. این به آن معناست که آنزیم فقط یکی از رشتههای DNA را خواهد شکست. به جای آن وقتی تمپلیتی از ترمیم همولوگ برای آن تدارک دیده شد، ترمیم DNA با حداکثر دقت انجام خواهد شد.
نوع سوم Cas9 نوکلئاز ناقص است. جهشهای ایجاد شده از برش DNA جلوگیری میکنند ولی توانایی اتصال به آن هنوز باقیست. بنابراین این تکنولوژی برای هدف قرار دادن توالیهای خاص بدون برش مفید خواهد بود. به جای آن با اتصال دامنههای عامل مختلف، این آنزیم میتواند برای خاموش یا فعال کردن ژنهای مختلف کاربرد داشته باشد. علاوه بر این، از نوع سوم آنزیم Cas9 میتوان برای تصویرسازی هم استفاده کرد؛ برای مثال از این مورد برای بالا بردن کیفیت استفاده از پروتئین فلوئورسنت سبز استفاده شدهاست.
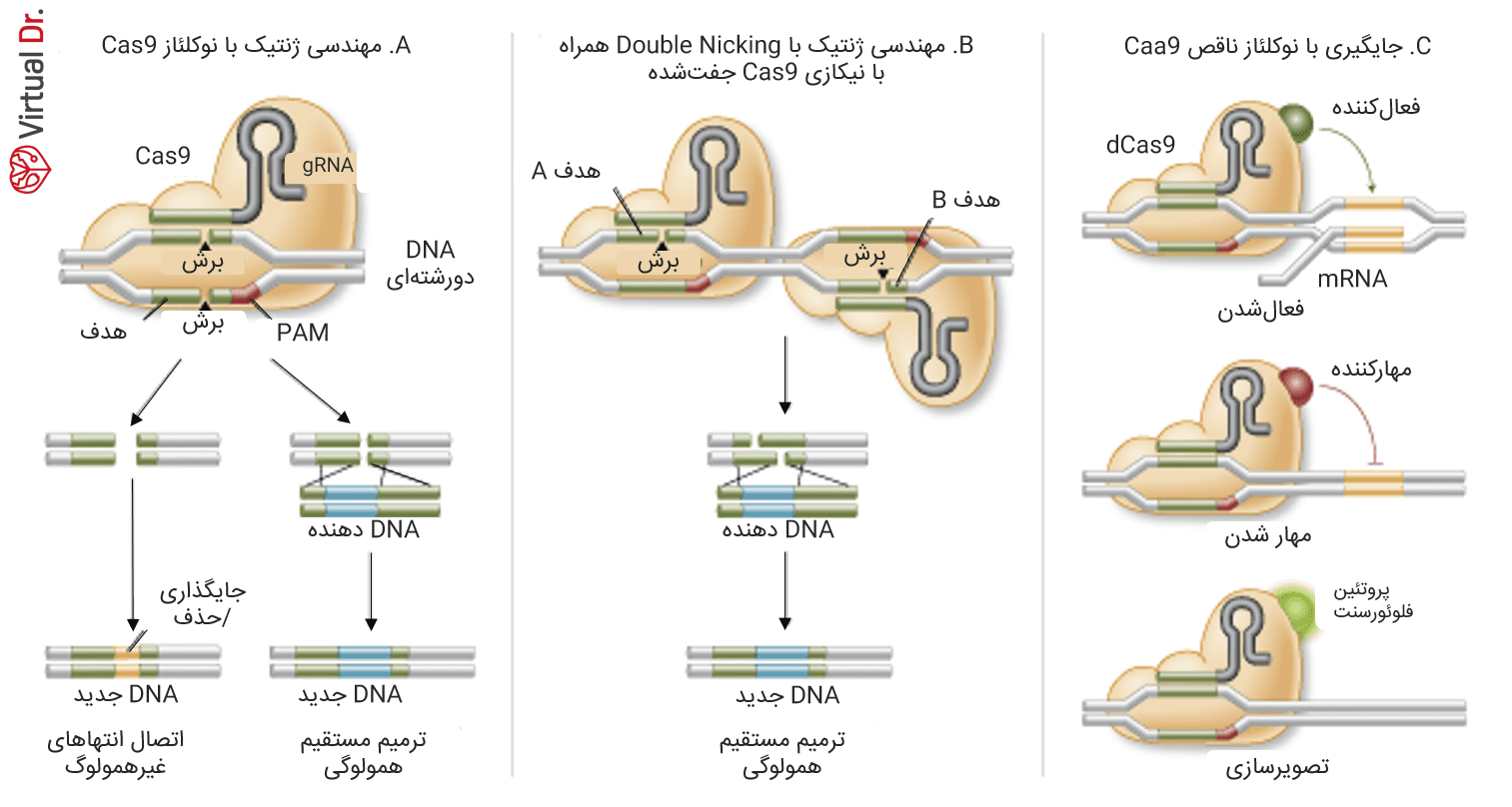
B) آنزیم Cas9 جهشیافته سایتی میسازد مخصوص برای نیک تک رشتهای. دو sgRNA میتواند برای برشی متقابل در دو رشته مقدمهای باشد، که میتواند متحمل ترمیم مستقیم همولوگی شود.
C) آنزیم Cas9 غیرنوکلئازی (نوع سوم) میتواند به دامنهیهای عامل زیادی متصل شده و جایگاههای مختلفی را ایجاد کند. برای مثال فعالکنندهها و مهارکنندههای رونویسی و همچنین پروتئینهای فلوئورسنت
کارآمدی هدفگیریها و جهشهای ناخواسته
کارایی هدفگیریها و درصد دستیابی به جهشهای دلخواه یکی از مهمترین پارامترها برای ارزیابی ابزار اصلاح ژنیست. مثلاً کارایی این روش در نوعی ماهی بالاتر از ۷۰% گزارش شده. این در حالیست که اثرات این سیستم در سلولهای بنیادی القایی ۲-۵% میباشد. از این سیستم تازه کشف شده به عنوان انقلابی در زمینهی بیولوژی سلولی مولکولی یاد میشود.
خیلی خوب بود. من دارم درمورد این تکنیک تحقیق میکنم و مطلبتون خیلی به دردم خورد مخصوصا فیلم انتهایی خیلی متن رو برام بازتر کرد. بی نهایت ممنون. سایت فوق العاده ای دارید. عالیه